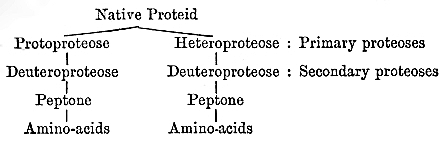
Native proteid, Protoproteose, Deuteroproteose, Heteroproteose, Primary proteoses, Secondary proteoses, Peptone, Amino-acids
Title: The nutrition of man
Author: R. H. Chittenden
Release date: August 24, 2022 [eBook #68830]
Language: English
Credits: Thiers Halliwell, Mark C. Orton and the Online Distributed Proofreading Team at https://www.pgdp.net (This file was produced from images generously made available by The Internet Archive)
The text of this e-book has been preserved in its original form apart from correction of the typographic errors listed below. Illustrations have been repositioned adjacent to relevant tabulated data, and the List of Illustrations adjusted accordingly. On p.72 an image of the Xanthin formula incorrectly shows a double bond between a carbon and nitrogen atom – the correct formula is shown on the next page – and there is a date discrepancy on p. 248 between the text and the illustration caption (November 18/February 27). Page numbers are shown in the right margin and footnotes are located at the end. Footnotes are located at the end.
Typographic corrections:
enyzmes → enzymes
oxgyen → oxygen
enyzme → enzyme
Futher → Further
mechancial → mechanical
rythmical → rhythmical
economcially → economically
circulirinden → circulirenden
SUBJECT → SUBJECTS
equibrium → equilibrium
availibility → availability
(166) grams → (166 grams)
accusstomed → accustomed
Glassner → Glässner
strach → starch
The present book is the outcome of a course of eight lectures delivered before the Lowell Institute of Boston in the early part of 1907.
In this presentation of the subject the attempt has been made to give a systematic account of our knowledge regarding some of the more important processes of nutrition, with special reference to the needs of the body for food. In doing this, the facts accumulated by painstaking observations and experiments during recent years in our laboratory have been incorporated with data from other sources and brought into harmony, so far as possible, with the modern trend of physiological thought.
Numerous experimental results, hitherto unpublished, have been introduced, notably in Chapter VII, in which a few of the data recently obtained in our laboratory with dogs are presented in some detail, since they afford evidence of the error of the current arguments concerning the necessity of a high proteid intake by man, as based on the results of earlier investigators with high proteid animals.
It is hoped that the facts and arguments here presented will help to arouse a more general interest in the subject of human nutrition, as right methods of living promise so much for the health and happiness of the individual and of the community.
vii
CHAPTER I | PAGE |
| Foods and their Digestion | |
| Topics: The purpose of nutrition. The food of man. Proteid foods. Carbohydrate foods. Fats. Food as fuel. Composition of foodstuffs. Availability of foods. Food as source of energy. Various factors in the nourishment of the body. Processes of digestion. Secretion of saliva. Function of saliva. Enzymes. Reversible action of enzymes. Specificity of enzymes. Mastication. Gastric secretion. Components of gastric juice. Action of gastric juice. Muscular movements of stomach. Time foods remain in stomach. Importance of stomach digestion. Processes of the small intestine. Secretion of pancreatic juice. Chemical changes in small intestine. Destruction of proteid food. Significance of the breaking down of proteid. Change of fatty foods and carbohydrates in intestine. Digestion practically complete at end of small intestine. Putrefaction held in check. Digestion a prelude to utilization of food. | |
CHAPTER II | |
| Absorption, Assimilation, and the Processes of Metabolism | |
| Topics: Physiological peculiarities in absorption. Chemical changes in epithelial walls of intestine. Two pathways for absorbed material. Function of the liver as a regulator of carbohydrate. Absorption of proteid products. Assimilation of food products. Anabolism. Katabolism. Metabolism. Processes of metabolism. Older views regarding oxidation. Discoveries of Lavoisier. The views of Liebig. Theory of luxus consumption. Oxidation in the body not simple combustion. Oxygen not the cause of the decompositions. Oxidation not confined to any one place. Intracellular enzymes. Living cells the guiding power in katabolism. Some intermediary products of tissue metabolism. Chemical structure of different proteids. Decomposition products of nucleoproteids. Relation to uric acid. Action of specific intracellular enzymes. Creatin and creatinin. Relation to urea. Proteid katabolism a series of progressive chemical decompositions. Intracellular enzymes as the active agents. | |
| viii CHAPTER III | |
| The Balance of Nutrition | |
| Topics: Body equilibrium. Nitrogen equilibrium. Carbon equilibrium. Loss of nitrogen during fasting. Influence of previous diet on loss of nitrogen in fasting. Output of carbon during fasting. Influence of pure proteid diet on output of nitrogen. Influence of fat on proteid metabolism. Effect of carbohydrate on nitrogen metabolism. Storing up of proteid by the body. Transformation of energy in the body. Respiration calorimeter. Basal energy exchange of the body. Circumstances influencing energy exchange. Effect of food on heat production. Respiratory quotient and its significance. Influence of muscle work on energy exchange. Elimination of carbon dioxide during work and with different diets. Effect of excessive muscular work on energy exchange. Oxygen consumption under different conditions. Output of matter and energy subject to great variation. Body equilibrium and approximate nitrogen balance to be expected in health. | |
CHAPTER IV | |
| Source of the Energy of Muscle Work, with Some Theories of Proteid Metabolism | |
| Topics: Relation of muscle work to energy exchange. Views of Liebig. Experimental evidence. Relation of nitrogen excretion to muscle work. Significance of the respiratory quotient in determining nature of the material oxidized. Fats and carbohydrates as source of energy by muscles. Utilization of proteid as a source of energy. Formation of carbohydrate from proteid. Significance of proteid metabolism. Theories of Carl Voit. Morphotic proteid. Circulating proteid. General conception of proteid metabolism on the basis of Voit’s theories. Pflüger’s views of proteid metabolism. Rapidity of elimination of food nitrogen. Methods by which nitrogen is split off from proteid. Theories of Folin. Significance of creatinin and of the percentage distribution of excreted nitrogen. Endogenous or tissue metabolism. Exogenous or intermediate metabolism. Needs of the body for proteid food possibly satisfied by quantity sufficient to meet the demands of tissue or endogenous metabolism. Bearings of Folin’s views on current theories and general facts of proteid metabolism. Large proteid reserve and voluminous exogenous metabolism probably not needed. Importance of feeding experiments in determining the true value of different views. | |
CHAPTER V | |
| Dietary Habits and True Food Requirements | |
| Topics: Dietetic customs of mankind. Origin of dietary standards. True food requirements. Arguments based on custom and habit. Relationshipix between food consumption and prosperity. Erroneous ideas regarding nutrition. Commercial success and national wealth not the result of liberal dietary habits. Instinct and craving not wise guides to follow in choice and quantity of food. Physiological requirements and dietary standards not to be based on habits and cravings. Old-time views regarding temperate use of food. The sayings of Thomas Cogan. The teachings of Cornaro. Experimental results obtained by various physiologists. Work of the writer on true proteid requirements. Studies with professional men. Nitrogen equilibrium with small amounts of food. Sample dietaries. Simplicity in diet. Nitrogen requirement per kilogram of body-weight. Fuel value of the daily food. Experiments with University athletes. Nitrogen balance and food consumption. Sample dietaries. Adequacy of a simple diet. | |
CHAPTER VI | |
| Further Experiments and Observations Bearing on True Food Requirements | |
| Topics: Dietary experiments with a detail of soldiers from the United States Army. General character of the army ration. Samples of the daily dietary adopted. Rate of nitrogen metabolism attained. Effect on body-weight. Nitrogen balance with lowered proteid consumption. Influence of low proteid on muscular strength of soldiers and athletes. Effect on fatigue. Effect on physical endurance. Fisher’s experiments on endurance. Dangers of underfeeding. Dietary observations on fruitarians. Observations on Japanese. Recent dietary changes in Japanese army and navy. Observations of Dr. Hunt on resistance of low proteid animals to poisons. Conclusions. | |
CHAPTER VII | |
| The Effect of Low Proteid Diet on High Proteid Animals | |
| Topics: A wide variety of foods quite consistent with temperance in diet. Safety of low proteid standards considered. Arguments based on the alleged effects of low proteid diet on high proteid animals. Experiments of Immanuel Munk with dogs. Experiments of Rosenheim. Experiments of Jägerroos. Comments on the above experiments. The experiments of Watson and Hunter on rats. The writer’s experiments with dogs. Details of the results obtained with six dogs. Comparison of the results with those of previous investigators. Effect of a purely vegetable diet on dogs. Different nutritive value of specific proteids considered. Possible influence of difference in chemical constitution of individual proteids. Effect of low proteid diet on the absorption and utilization of food materials in the intestine of dogs. General conclusions from the results of experiments with animals. | |
| x CHAPTER VIII | |
| Practical Applications with Some Additional Data | |
| Topics: Proper application of the results of scientific research helpful to mankind. Dietary habits should be brought into conformity with the true needs of the body. The peculiar position of proteid foods emphasized. The evil effects of overeating. What the new dietary standards really involve. The actual amounts of foodstuffs required. Relation of nutritive value to cost of foods. The advantages of simplicity in diet. A sample dietary for a man of 70 kilograms body-weight. A new method of indicating food values. Moderation in the daily dietary leads toward vegetable foods. The experiments of Dr. Neumann. The value of fruits as food. The merits of animal and vegetable proteids considered in relation to the bacterial processes in the intestine. A notable case of simplicity in diet. Intelligent modification of diet to the temporary needs of the body. Diet in summer and winter contrasted. Value of greater protection to the kidneys. Conclusion. | |
| INDEX |
xi
| FACING PAGE | |
| Photograph of one of the athletes | 190 |
| Photograph of soldiers taken at the close of the experiment | 194 |
| Photograph of soldiers taken at the close of the experiment | 194 |
| Photograph of Fritz at the close of the experiment | 200 |
| Photographs of the dogs experimented with | |
| Subject No. 5 August 19, 1905 | |
| Subject No. 5 November 18, 1905 | |
| Subject No. 5 April 24, 1906 | |
| Subject No. 5 June 27, 1906 | |
| Subject No. 3 August 19, 1905 | |
| Subject No. 3 November 18, 1905 | |
| Subject No. 3 April 24, 1906 | |
| Subject No. 3 June 27, 1906 | |
| Subject No. 13 January 2, 1906 | |
| Subject No. 13 February 27,1906 | |
| Subject No. 13 April 24, 1906 | |
| Subject No. 13 June 19, 1906 | |
| Subject No. 15 January 2, 1906 | |
| Subject No. 15 February 27, 1906 | |
| Subject No. 15 April 24, 1906 | |
| Subject No. 15 June 19, 1906 | |
| Subject No. 20 January 2, 1906 | |
| Subject No. 20 February 27, 1906 | |
| Subject No. 20 April 24, 1906 | |
| Subject No. 20 June 19, 1906 | |
| Subject No. 17 January 2, 1906 | |
| Subject No. 17 February 27, 1906 | |
| Subject No. 17 April 24, 1906 | |
| Subject No. 17 June 27, 1906 |
1
FOODS AND THEIR DIGESTION
Topics: The purpose of nutrition. The food of man. Proteid foods. Carbohydrate foods. Fats. Food as fuel. Composition of foodstuffs. Availability of foods. Food as source of energy. Various factors in the nourishment of the body. Processes of digestion. Secretion of saliva. Function of saliva. Enzymes. Reversible action of enzymes. Specificity of enzymes. Mastication. Gastric secretion. Components of gastric juice. Action of gastric juice. Muscular movements of stomach. Time foods remain in stomach. Importance of stomach digestion. Processes of the small intestine. Secretion of pancreatic juice. Chemical changes in small intestine. Destruction of proteid food. Significance of the breaking down of proteid. Change of fatty foods and carbohydrates in intestine. Digestion practically complete at end of small intestine. Putrefaction held in check. Digestion a prelude to utilization of food.
One of the great mysteries of life is the power of growth, that harmonious development of composite organs and tissues from simple protoplasmic cells, with the ultimate formation of a complex organism with its orderly adjustment of structure and function. Equally mysterious is that wonderful power of rehabilitation by which the cells of the body are able to renew their living substance and to maintain their ceaseless activity through a period, it may be of fourscore years, before succumbing to the inevitable fate that awaits all organic structures. This bodily activity, visible and invisible, is the result of a third mysterious process, more or less continuous as long as life endures, of chemical2 disintegration, decomposition, and oxidation, by which arises the evolution of energy to maintain the heat of the body and the power for mental and physical work.
These three main functions constitute the purpose of nutrition. The growth of the adult man from the tiny cell or germ that marks his simple beginning is at the expense of the food material he absorbs and assimilates. The rehabilitation of the cells, or the composite tissues of the fully developed organism, is accomplished through utilization of the daily food, whereby cell substance is renewed and all losses made good. The energy which manifests itself in the form of heat and mechanical or mental work, i. e., the energy by which the vital machinery is maintained in ceaseless activity, comes from the breaking down of the food materials by means of which, as the saying goes, the body is nourished. The body thus becomes the centre of different lines of activity, the food serving as the material out of which new cells and tissues are constructed, old cells revivified, and energy for running the bodily machinery derived. Development, growth, and vital activity all depend upon the availability of food in proper amounts and proper quality.
The food of man is composed mainly of organic materials, for while, as Dr. Curtis1 has expressed it, “the plant can make organic matter out of inorganic elements, just this the animal cannot do at all. The thing of legs and locomotion, of spine and speech, can build his organic walls only out of organic bricks ruthlessly ripped from existing walls of other animals or plants.” It is true that man has need of certain inorganic salts in his daily diet, but they are in the nature of aids to nutrition (aside from such as are necessary for the formation of bone and teeth), contributing in some measure toward regulation and control of nutritive processes rather3 than as a source of energy to the body. Inorganic substances, however, are an integral part of the essential tissues and organs of the body, being combined with the organic constituents of the living cells. Indeed, electrolytes are perhaps the substances that put life into the proteids of the protoplasm, and it is truly important for the integrity and functional power of living cells that the proportion of inorganic constituents therein be kept in a constant condition of quality and quantity. Still, the food of mankind is essentially organic in nature, and while it may be exceedingly varied in character, ranging from the simple vegetable dietary of the natives of India and the Far East to the voluminous admixture of varied forms of animal and vegetable foodstuffs so acceptable to the bon vivant of our western civilization, the principles contained therein are few in number.
The organic foodstuffs are of three distinct types and are classified under three heads, viz.: Proteids or Albuminous foodstuffs, Carbohydrates, and Fats. All animal and vegetable foods, whatever their nature and whatever their origin, are composed simply of representatives of one or more of these three classes of food principles.
Proteid substances are characterized by containing about 16 per cent of nitrogen. In addition, they contain on an average 52 per cent of carbon, 7 per cent of hydrogen, 23 per cent of oxygen, and 0.5–2.0 per cent of sulphur. A certain class of proteids, known as nucleoproteids because of their occurrence in the nuclei of cells, contain likewise a small amount of phosphorus in organic combination. Proteid or albuminous substances constitute the chemical basis of all living cells, whether animal or vegetable. This means, expressed in different language, that the organic substance of all organs and tissues, whether of animals or plants, is made up principally of proteid matter. Proteid substances occupy, therefore, a peculiar position in the nutrition of man4 and of animals in general. They constitute the class of essential foodstuffs without which life is impossible. For tissue-building and for the renewal of tissues and organs, or their component cells, proteid or albuminous foodstuffs are an absolute requirement. The vital part of all tissue is proteid, and only proteid food can serve for its growth or renewal. Hence, no matter how generous the supply of carbohydrates and fats, without some admixture of proteid food the body will weaken and undergo “nitrogen starvation.” It is to be noted, however, that while the element nitrogen (16 per cent) gives character to the proteid or albuminous foodstuffs, so that they are frequently spoken of or classified as the “nitrogenous foodstuffs,” it is not the nitrogen per se that is so essential for the nutrition of the body. Man lives in an atmosphere of oxygen and nitrogen. He can and does absorb and utilize the free oxygen of the air he breathes; indeed, it is absolutely essential for his existence, but the free nitrogen likewise drawn into the lungs at each inspiration is of no avail for the needs of the body. Further, there are many compounds of nitrogen, some of them closely allied to the proteid foodstuffs in chemical composition, which are just as useless as free nitrogen in meeting the wants of the body for nitrogenous foods.
Dame Nature is very discriminating; she demands a definite form of nitrogenous compound, some peculiar or specific grouping of the nitrogen element with other elements in the food that can make good the waste of proteid tissue. In the inactive and fibrous tissues of animals, such as are found in bones, tendons, and ligaments, there is present a substance known as collagen, which, when boiled with water, as in the making of soups, is transformed into gelatin. This body, because of its close chemical relationship to proteid or albuminous substances, is known as an albuminoid. Yet, though it has essentially the same chemical composition as ordinary5 albuminous substances and shows many of the reactions characteristic of the latter, it cannot take the place of true proteid in building up or repairing the tissues of the body. To quote again from Dr. Curtis: “Tissue is nitrogenous, so that, of course, only nitrogenous food can serve for its making; but of the two kinds of nitrogenous principles, proteids and albuminoids, behold, proteids only are of avail! Why this is so is unknown, since albuminoid is equally nitrogenous with proteid; but so it is—proteid and proteid alone can fulfil the high function of furnishing the material basis of life. Gelatin cannot even go to make the very kind of tissue of which itself is a derivative. Alongside of its brother proteid, gelatin stands as a prince of the blood whose escutcheon bears the ‘bend sinister.’ Such a one, though of royal lineage, may never aspire to the throne.” It is thus quite clear that the true proteid foods are tissue builders in the broadest sense of the term, and it is equally evident that they are absolutely essential for life, since no other kind or form of foodstuff can take their place in supplying the needs of the body. Every living cell, whether of heart, muscle, brain, or nerve, requires its due allowance of proteid material to maintain its physiological rhythm. No other foodstuff stands in such intimate relationship to the vital processes, but so far as we know at present any form of true proteid, whether animal or vegetable, will serve the purpose.
Carbohydrates include two closely related classes of compounds, viz., sugars and starches. They are entirely free from nitrogen, containing only carbon (44.4 per cent), hydrogen (6.2 per cent), and oxygen (49.4 per cent), and hence are classified as non-nitrogenous foods. Obviously, they cannot serve as tissue builders, but by oxidation they yield energy for heat and work. They constitute an easily oxidizable form of fuel, and when supplied in undue amounts they may undergo transformation within the body into fat, which6 is temporarily deposited in tissues and organs for future needs.
Fats, like carbohydrates, are free from nitrogen, but differ from them in containing a much larger percentage of carbon, and hence have greater fuel value per pound. Fats contain on an average 76.5 per cent of carbon, 11.9 per cent of hydrogen, and 11.5 per cent of oxygen. With their larger content of carbon and smaller proportion of oxygen, fats are less easily oxidizable than sugars, requiring a larger intake of oxygen for their combustion, but when oxidized they yield more heat per pound than carbohydrates.
Fats and carbohydrates are thus seen to be the natural fuel foodstuffs of the body. They cannot serve for the upbuilding or renewal of tissue, but by oxidation they constitute an economical fuel for maintaining body temperature and for power to run the bodily machinery. It should be remembered, however, that anything capable of being burned in the body may serve as fuel material; hence proteid food, though of specific value as a tissue builder, may likewise by its oxidation yield energy for heat and work, but its combustion, owing to the content of nitrogen, is never complete. Further, its use as fuel is uneconomical and undesirable for reasons to be discussed later, but it is well to know that its oxidation, though incomplete, is accompanied by the liberation of energy, as in the oxidation of non-nitrogenous foods. A portion of the carbon, hydrogen, and oxygen of the proteid molecule will burn within the body to gaseous products, as do sugars and fats, but there remains a nucleus of nitrogen, with some carbon, hydrogen, and oxygen, which resists combustion and must be gotten rid of by the combined labors of liver and kidneys. Fats and carbohydrates, on the other hand, undergo complete combustion to simple gaseous products, carbon dioxide and water, which are easily removed by the lungs, skin, etc.
7
These three classes of foodstuffs exist in a great variety of combinations or admixtures in nature. In many cases, noticeably in milk, all three occur together in fairly large quantities. In animal foods, such as meats, fish, etc., proteid and fat alone are found, while in perfectly lean meat proteid only is present, excepting a small amount of fat. Again, the white of the egg contains proteid alone. Hence, a meat and egg diet would be essentially a proteid diet. In vegetable foods, as in the cereals, there is found an admixture of proteid and starch, the latter predominating in many cases, as in wheat flour. The following table,2 showing the chemical composition of various food materials, may be of service in throwing light on the relative distribution of the three classes of foodstuffs in natural products.
THE CHEMICAL COMPOSITION OF SOME COMMON FOOD MATERIALS
Food Materials. |
Proteid. |
Carbo-hydrate. |
Fat. |
Water. |
Mineral Matter. |
Fuel Value per pound. |
|---|---|---|---|---|---|---|
per cent |
per cent |
per cent |
per cent |
per cent |
calories |
|
| Fresh beef, loin, lean, edible portion | 24.2 |
0 |
3.7 |
70.8 |
1.3 |
615 |
| Fresh beef, round, lean, edible portion | 22.3 |
0 |
2.8 |
73.6 |
1.3 |
540 |
| Fresh Porterhouse steak, edible portion | 21.9 |
0 |
20.4 |
60.0 |
1.0 |
1270 |
| Fresh beef liver | 21.0 |
1.7 |
4.5 |
71.2 |
1.6 |
605 |
| Fresh beef tongue | 19.0 |
0 |
9.2 |
70.8 |
1.0 |
740 |
| Fresh sweetbreads | 16.8 |
0 |
12.1 |
70.9 |
1.6 |
825 |
| Fresh beef kidney | 16.9 |
0.4 |
4.8 |
76.7 |
1.2 |
520 |
| Cooked beef, roasted | 22.3 |
0 |
28.6 |
48.2 |
1.3 |
1620 |
| Cooked round steak | 27.6 |
0 |
7.7 |
63.0 |
1.8 |
840 8 |
| Broiled tenderloin steak | 23.5 |
0 |
20.4 |
54.8 |
1.2 |
1300 |
| Dried beef, canned | 39.2 |
0 |
5.4 |
44.8 |
11.2 |
960 |
| Stewed kidneys, canned | 18.4 |
2.1 |
5.1 |
71.9 |
2.5 |
600 |
| Fresh corned beef, edible portion | 15.3 |
0 |
26.2 |
53.6 |
4.9 |
1395 |
| Fresh breast of veal, lean | 21.2 |
0 |
8.0 |
70.3 |
1.0 |
730 |
| Fresh leg of lamb, edible portion | 19.2 |
0 |
16.5 |
63.9 |
1.1 |
1055 |
| Lamb chops, broiled | 21.7 |
0 |
29.9 |
47.6 |
1.3 |
1665 |
| Roast leg of lamb, edible portion | 19.4 |
0 |
12.7 |
67.1 |
0.8 |
900 |
| Roast leg of mutton, edible portion | 25.9 |
0 |
22.6 |
50.9 |
1.2 |
1420 |
| Fresh lean ham | 25.0 |
0 |
14.4 |
60.0 |
1.3 |
1075 |
| Smoked ham, fat, edible portion | 14.8 |
0 |
52.3 |
27.9 |
3.7 |
2485 |
| Chicken, broilers, edible portion | 21.5 |
0 |
2.5 |
74.8 |
1.1 |
505 |
| Turkey, edible portion | 21.1 |
0 |
22.9 |
55.5 |
1.0 |
1360 |
| Roast turkey, edible portion | 27.8 |
0 |
18.4 |
52.0 |
1.2 |
1295 |
| Fricasseed chicken, edible portion | 17.6 |
2.4 |
11.5 |
67.5 |
1.0 |
855 |
| Fresh cod, dressed | 11.1 |
0 |
0.2 |
58.5 |
0.8 |
215 |
| Fresh mackerel, edible portion | 18.7 |
0 |
7.1 |
73.4 |
1.2 |
645 |
| Fresh halibut, steaks | 18.6 |
0 |
5.2 |
75.4 |
1.0 |
565 |
| Fresh shad, edible portion | 18.8 |
0 |
9.5 |
70.6 |
1.3 |
750 |
| Fresh smelt, edible portion | 17.6 |
0 |
1.8 |
79.2 |
1.7 |
405 |
| Cooked bluefish, edible portion | 26.1 |
0 |
4.5 |
68.2 |
1.2 |
670 |
| Broiled Spanish mackerel, edible portion | 23.2 |
0 |
6.5 |
68.9 |
1.4 |
715 9 |
| Salt codfish, edible portion | 25.4 |
0 |
0.3 |
53.5 |
24.7 |
410 |
| Salt mackerel, edible portion | 22.0 |
0 |
22.6 |
42.2 |
13.2 |
1345 |
| Canned salmon, edible portion | 21.8 |
0 |
12.1 |
63.5 |
2.6 |
915 |
| Canned sardines, edible portion | 23.0 |
0 |
19.7 |
52.3 |
5.6 |
162 |
| Fresh round clams | 6.5 |
4.2 |
0.4 |
86.2 |
2.7 |
215 |
| Fresh oysters, solid | 6.0 |
3.3 |
1.3 |
88.3 |
1.1 |
230 |
| Fresh hen’s eggs | 13.4 |
0 |
10.5 |
73.7 |
1.0 |
720 |
| Boiled hen’s eggs | 13.2 |
0 |
12.0 |
73.2 |
0.8 |
765 |
| Butter | 1.0 |
0 |
85.0 |
11.0 |
3.0 |
3605 |
| Full cream cheese | 25.9 |
2.4 |
33.7 |
34.2 |
3.8 |
1950 |
| Whole cow’s milk | 3.3 |
5.0 |
4.0 |
87.0 |
0.7 |
325 |
| Corn meal, unbolted | 8.4 |
74.0 |
4.7 |
11.6 |
1.3 |
1730 |
| Oatmeal | 16.1 |
67.5 |
7.2 |
7.3 |
1.9 |
1860 |
| Rice | 8.0 |
79.0 |
0.3 |
12.3 |
0.4 |
1630 |
| Wheat flour, entire wheat | 13.8 |
71.9 |
1.9 |
11.4 |
1.0 |
1675 |
| Boiled rice | 2.8 |
24.4 |
0.1 |
72.5 |
0.2 |
525 |
| Shredded wheat | 10.5 |
77.9 |
1.4 |
8.1 |
2.1 |
1700 |
| Macaroni | 13.4 |
74.1 |
0.9 |
10.3 |
1.3 |
1665 |
| Brown bread | 5.4 |
47.1 |
1.8 |
43.6 |
2.1 |
1050 |
| Wheat bread or rolls | 8.9 |
56.7 |
4.1 |
29.2 |
1.1 |
1395 |
| Whole wheat bread | 9.4 |
49.7 |
0.9 |
38.4 |
1.3 |
1140 |
| Soda crackers | 9.8 |
73.1 |
9.1 |
5.9 |
2.1 |
1925 |
| Oyster crackers | 11.3 |
70.5 |
10.5 |
4.8 |
2.9 |
1965 |
| Ginger bread | 5.8 |
63.5 |
9.0 |
18.8 |
2.9 |
1670 |
| Sponge cake | 6.3 |
65.9 |
10.7 |
15.3 |
1.8 |
1795 |
| Lady fingers | 8.8 |
70.6 |
5.0 |
15.0 |
0.6 |
1685 |
| Apple pie | 3.1 |
42.8 |
9.8 |
42.5 |
1.8 |
1270 |
| Custard pie | 4.2 |
26.1 |
6.3 |
62.4 |
1.0 |
830 |
| Squash pie | 4.4 |
21.7 |
8.4 |
64.2 |
1.3 |
840 10 |
| Indian meal pudding | 5.5 |
27.5 |
4.8 |
60.7 |
1.5 |
815 |
| Tapioca pudding | 3.3 |
28.2 |
3.2 |
64.5 |
0.8 |
720 |
| Fresh asparagus | 1.8 |
3.3 |
0.2 |
94.0 |
0.7 |
105 |
| Fresh lima beans | 7.1 |
22.0 |
0.7 |
68.5 |
1.7 |
570 |
| Dried lima beans | 18.1 |
65.9 |
1.5 |
10.4 |
4.1 |
1625 |
| Dried beans | 22.5 |
59.6 |
1.8 |
12.6 |
3.5 |
1605 |
| Cooked beets | 2.3 |
7.4 |
0.1 |
88.6 |
1.6 |
185 |
| Fresh cabbage, edible portion | 1.6 |
5.6 |
0.3 |
91.5 |
1.0 |
145 |
| Green corn, edible portion | 3.1 |
19.7 |
1.1 |
75.4 |
0.7 |
470 |
| Dried peas | 24.6 |
62.0 |
1.0 |
9.5 |
2.9 |
1655 |
| Green peas | 7.7 |
16.9 |
0.5 |
74.6 |
1.0 |
465 |
| Raw potatoes, edible portion | 2.2 |
18.4 |
0.1 |
78.3 |
1.0 |
385 |
| Boiled potatoes | 2.5 |
20.9 |
0.1 |
75.5 |
1.0 |
440 |
| Fresh tomatoes | 0.9 |
3.9 |
0.4 |
94.3 |
0.5 |
105 |
| Baked beans, canned | 6.9 |
19.6 |
2.5 |
68.9 |
2.1 |
600 |
| Apples, edible portion | 0.4 |
14.2 |
0.5 |
84.6 |
3.0 |
290 |
| Bananas, yellow, edible portion | 1.3 |
22.0 |
0.6 |
75.3 |
0.8 |
460 |
| Fresh cranberries | 0.4 |
9.9 |
0.6 |
88.9 |
0.2 |
215 |
| Oranges, edible portion | 0.8 |
11.6 |
0.2 |
86.9 |
0.5 |
240 |
| Peaches, edible portion | 0.7 |
9.4 |
0.1 |
89.4 |
0.4 |
190 |
| Fresh strawberries | 1.0 |
7.4 |
0.6 |
90.4 |
0.6 |
180 |
| Dried prunes, edible portion | 2.1 |
73.3 |
0.0 |
22.3 |
2.3 |
1400 |
| Almonds, edible portion | 21.0 |
17.3 |
54.9 |
4.8 |
2.0 |
3030 |
| Peanuts, edible portion | 25.8 |
24.4 |
38.6 |
9.2 |
2.0 |
2560 |
| Pine nuts, edible portion | 33.9 |
6.9 |
49.4 |
6.4 |
3.4 |
2845 |
| Brazil nuts, edible portion | 17.0 |
7.0 |
66.8 |
5.3 |
3.9 |
3265 |
| Soft-shell walnuts, edible portion | 16.6 |
16.1 |
63.4 |
2.5 |
1.4 |
3285 11 |
In commenting on these figures, reference to which will be made from time to time in other connections, it may be wise to emphasize the large amount of water almost invariably present in natural foodstuffs. Further, it is to be noted that, in animal products especially, the variations in proteid-content are in large measure coincident with variations in the amount of water present. In other words, foods of animal origin if freed entirely of water would, as a rule, show essentially the same percentage of proteid matter. Fat is naturally variable, according to the condition of the animal at the time it was slaughtered. Among the vegetable products, carbohydrate, mainly in the form of starch, becomes exceedingly conspicuous, though proteid is by no means lacking. Indeed, in some cereals, as in oatmeal, in dried peas and beans, the content of proteid will average as high as in fresh beef, while in addition 50–70 per cent of the entire substance is made up of carbohydrate. Again, in the edible nuts, the content of proteid runs high, in some cases higher than in fresh beef, while at the same time carbohydrate and fat are noticeably large. Further, it is to be noted that in nuts there is here and there some striking individuality, as in pine nuts and Brazil nuts, both of which show a noticeable lack of carbohydrate as contrasted with peanuts, almonds, and walnuts; a fact of some importance in cases where a vegetable food rich in proteid is desired, but with freedom from starch.
Another generality, to be thoroughly understood, is that while the figures given for proteid express quite clearly and with reasonable degree of accuracy the relative amounts of proteid matter present in the foodstuffs in question, there may be important differences in availability of which the percentage figures give no suggestion. In other words, the analytical data deal solely with the total content of proteid, while there is needed in addition information as to the relative digestibility, or availability by the body, of the different12 kinds of proteid food. For example, roast mutton, cream cheese, and dried peas contain approximately the same amount of proteid. Are we then to infer that these three foods have the same nutritive value so far as proteid is concerned? Surely not, since no account is taken of the relative digestibility of the three foods. It is one of the axioms of physiology that the true nutritive value of any proteid food is dependent not alone upon the amount of proteid contained therein, but upon the quantity of proteid that can be digested and absorbed; or, in other words, made available for the needs of the body. The same rule holds good for both fats and carbohydrates, but as proteid is the more important foodstuff, and is as a rule taken more sparingly, the question of availability has greater import with the proteid foods.
The availability or digestibility of foods can be determined only by physiological experiment. By making a comparison for a definite period of time of the amount of a given food ingredient consumed and the amount that passes unchanged through the intestine, an estimate of its digestibility can be made. The result, to be sure, is not wholly free from error, since we cannot always distinguish between the undigested food and so-called metabolic products coming from the digestive juices and from the walls of the intestine; but the errors are not large, and results so obtained are full of meaning. In a general way it may be stated that with animal foods, such as meats, eggs, and milk, about 97 per cent of the contained proteid is digested and thereby rendered available for the body. With ordinary vegetable foods, on the other hand, as they are usually prepared for consumption, only about 85 per cent of the proteid is made available. This is partially due to the presence in the vegetable tissue of cellulose, which in some measure prevents that thorough attack of the proteid by the digestive juices which occurs with animal13 foods. With a mixed diet, i. e., with a variable admixture of animal and vegetable foods, it is usually considered that about 92 per cent of the proteid contained therein will undergo digestion.
Regarding differences in the availability of fats, it may be stated that, as a rule, the fatty matter contained in vegetable foods is less readily, or less thoroughly, digested than that present in foods of animal origin. In the latter, about 95 per cent of the fat is digested and absorbed. This figure, however, is generally taken as representing approximately the digestibility or availability of the fat contained in man’s daily dietary, since by far the larger proportion of the fat consumed is of animal origin. Carbohydrates, on the other hand, are much more easily utilized by the body. Naturally, sugars, owing to their great solubility and ready diffusibility, offer little difficulty in the way of easy digestion; but starches likewise, though not so readily assimilable, are digested, as a rule, to the extent of 98 per cent or more of the amount consumed. It is thus evident that in any estimate of the food value of a given diet, chemical composition is to be checked by the digestibility or availability of the food ingredients.
As has been stated several times, the proteid foodstuffs are the more important, since proteid matter is essential to animal life. Man must have a certain amount of proteid food to maintain the body in a condition of strength and vigor. The other essential is that the daily food furnish sufficient energy to meet the needs of the body for heat and power. This means that in addition to proteid, which primarily serves a particular purpose, there must be enough non-nitrogenous food (either carbohydrate or fat or both) to provide the requisite fuel for oxidation or combustion to meet the demands of the body for heat and for work; both of which are subject to great variation owing to differences14 in the temperature of the surrounding air, and especially because of variations in the degree of bodily activity. The energy which a given foodstuff will yield can be ascertained by laboratory experiment, in which a definite weight of the substance is burned or oxidized in a calorimetric bomb under conditions where the exact amount of heat liberated can be accurately measured. The fuel, or energy, value so obtained is expressed in calories or heat units. A calorie may be defined as the amount of heat required to raise 1 gram of water 1° C., or, to be more exact, the amount of heat required to raise 1 gram of water from 15° to 16° C. This unit is usually spoken of as the small calorie, to distinguish it from the large calorie, which represents the amount of heat required to raise 1 kilogram of water 1° C. Hence, the large calorie is equal to one thousand small calories. When burned in a calorimeter, 1 gram of carbohydrate yields on an average 4100 gram-degree units of heat, or small calories; 1 gram of fat yields 9300 small calories. Both of these non-nitrogenous foods burn or oxidize to the same products—viz., carbon dioxide and water—when utilized in the body as when burned in the calorimeter; hence, the figures given represent the physiological heat of combustion, per gram, of the two classes of foodstuffs. Obviously, the fuel values of different foods belonging to the same group or class will show slight variation, but the above figures represent average values.
Unlike fats and carbohydrates, proteids are not burned completely in the body; hence, the physiological fuel value of a proteid is less than the value obtained by oxidation in a bomb calorimeter. In the body, proteids yield certain decomposition products which are removed through the excreta, and which represent a certain quantity of potential energy thus lost to the economy. The average fuel value of proteids burned outside of the body is placed at 5711 calories15 per gram,3 or 5.7 large calories. Deducting the heat value of the proteid decomposition products contained in the excreta, the physiological fuel value of proteids is reduced on an average to about 4.1 large calories per gram.4 Rubner considers that the physiological fuel value of vegetable proteids is somewhat less than that of animal proteids; conglutin, for example, yielding 3.96 calories, as contrasted with 4.3 calories furnished by egg-albumin, or 4.40 calories from casein. On a mixed diet, where 60 per cent of the ingested proteid food is of animal origin and 40 per cent vegetable, the fuel value available to the body would be about 4.1 calories per gram of proteid, on the assumption that the physiological heat value of vegetable proteids averages 3.96 calories per gram and that of animal proteids 4.23 calories per gram (Rubner).
At present, we accept for all purposes of computation the following figures as representing the physiological or available (to the body) fuel value of the three classes of organic foodstuffs:
| 1 gram of proteid | 4.1 Large Calories |
| 1 gram of fat | 9.3 " " |
| 1 gram of carbohydrate | 4.1 " " |
From these data, it is evident at a glance that 1 gram of fat is isodynamic with 2.27 grams of either carbohydrate or proteid; and since carbohydrate and fat are of use to the body mainly because of their energy value, it is obvious that 50 grams of fat taken as food will be of as much service to the body as 113 grams of starch. In view of the relatively high fuel value of fats, it follows that the physiological heat of combustion of any given food material will correspond16 largely with the content of fat therein. This is quite apparent from the data given in the table showing chemical composition of food materials, where the fuel value per pound is seen to run more or less closely parallel with the percentage of fat. Experience, as well as direct physiological experiment, teaches us, however, that fat and carbohydrate cannot be interchanged indefinitely, because of the difficulty in utilization of fat when the amount is increased beyond a certain point. Personal experience provides ample evidence of the difference in availability between the two classes of foodstuffs. Carbohydrates are easily utilizable, fats with more difficulty. Palate, as well as stomach, rebels at large quantities of fat; a statement that certainly holds good for most civilized people, though exceptions may be found, as in the Esquimeaux and certain savage races.
In the nourishment of the body, the various factors that aid in the utilization of food are of great moment and must not be overlooked. It is not enough that the body be supplied with the proper proportion of nutrients, with sufficient proteid to meet the demand for nitrogen, and with carbohydrate and fat adequate to yield the needed energy; but all those physiological processes which have to do with the preparation of the foodstuffs for absorption into the circulating blood and lymph must be in effective working order. There is an intricacy of detail here which calls for careful oversight, and it is one of the functions of the nervous system to control and regulate both the mechanical and the chemical processes that are concerned in this seemingly automatic progression of foodstuffs from their entry into the mouth cavity to their final discharge from the alimentary tract, after removal of the last vestige of true nutritive material.
Mastication; deglutition; secretion of the various digestive juices, saliva, gastric juice, pancreatic juice, bile, intestinal juice, etc.; peristalsis, or the rhythmical movements of the17 muscular walls of the gastro-intestinal tract; the solvent action of the several digestive fluids on the different types of foodstuffs; the absorption of the products formed as a preliminary step in their transportation to the tissues and organs of the body, where they are to serve their ultimate purpose in nutrition; the interaction of these several processes one on the other; and, finally, the influence of the various nerve fibres and nerve centres concerned in the control of these varied activities,—all must work together in harmony and precision if the full measure of available nitrogen and energy-yielding material is to be extracted and absorbed from the ingested food, without undue expenditure of physiological labor. Further, the various processes of cell and tissue metabolism, by which the absorbed food material is built up into living protoplasm, and the chemical processes of oxidation, hydrolysis, reduction, etc., by which the intra and extra cellular material is broken down progressively into varied katabolic or excretory products, with liberation of energy; all these must move forward harmoniously and with due regard to the preservation of an even balance between intake and outgo, if the nutrition of the body is to be maintained at a proper level, and with that degree of physiological economy which is coincident with good health and high efficiency.
We may well pause here and consider briefly some of these processes which play so prominent a part in the proper utilization of the three classes of organic foodstuffs. The first digestive fluid which the ingested food comes in contact with is the saliva. Sensory nerve fibres, chiefly of the glossopharyngeal and lingual nerves which supply the mouth and tongue, are stimulated by the sapid substances of the food, and likewise by mere contact of the food particles with the mucous membrane lining the mouth cavity as the food is masticated and rolled about prior to deglutition. Impulses18 communicated in this way to the above sensory nerves are transmitted to certain nerve centres in the medulla oblongata, whence impulses are reflected back through secretory nerves going to the individual salivary glands, thereby calling forth a secretion. The production of saliva is thus a simple reflex act, in which the food consumed serves as a true stimulant or excitant. Pawlow,5 indeed, claims a certain degree of adaptability of the secretion to the character of the food taken into the mouth. Thus, he finds that dry, solid food excites a large flow of saliva, such as would be needed to masticate it properly and bring it into a suitable condition for swallowing. On the other hand, foods containing an abundance of water cause only a scanty flow of saliva. The situation of this secretory centre in the medulla, and the many branchings of nerve cells in this locality would naturally suggest the possibility of salivary secretion being incited by stimuli from a variety of sources. This is indeed the case, and it is worthy of note that a flow of saliva may result from stimulation of the sensory fibres of the vagus nerves as well as of the splanchnic and sciatic, thus indicating how a given secreting gland may be called into activity by impulses or stimuli which come to the centre through very indirect and devious pathways. Further, the secretory centre may be stimulated, and likewise inhibited, by impulses which have their origin in higher nerve centres in the brain. These facts are of great importance in throwing light upon the ways in which a secretion like saliva is called forth and its digestive action thus made possible. The thought and the odor of savory food cause the mouth to water, the flow of saliva so incited being the result of psychical stimulation. Similarly, fear, embarrassment, and anxiety frequently cause a dry mouth and parched throat through inhibition of the19 secretory centre by impulses which have their origin in higher centres in the brain.
The application of these facts to our subject is perfectly obvious, since they suggest at once how the production or secretion of an important digestive fluid—upon which the utilization of a given class of foodstuffs may be quite dependent—is controlled and modified through the nervous system by a variety of circumstances. We might reason that the appearance, odor, and palatability of food are factors of prime importance in its utilization by the body; that the æsthetics of eating are not to be ignored, since they have an important influence upon the flow of the digestive secretions. A peaceful mind, pleasurable anticipation, freedom from care and anxiety, cheerful companionship, all form desirable table accessories which play the part of true psychical stimuli in accelerating the flow of the digestive juices and thus pave the way for easy and thorough digestion. Further, it is easy to see how thorough mastication of food may prolong mechanical stimulation of the salivary glands and thus increase the flow of the secretion, while the longer stay of sapid substances in the mouth cavity increases the duration of the chemical stimulation of the sensory fibres of the lingual and glossopharyngeal nerves. In this connection, we may cite the view recently advanced by Pawlow that the individual salivary glands respond normally to different stimuli. Thus, there are three pairs of salivary glands concerned in the production of saliva,—the submaxillary, parotid, and sublingual,—all of which pour their secretions through separate ducts into the mouth cavity. By experiment, Pawlow has found that in the dog the submaxillary gland yields a copious flow of saliva when stimulated by acids, the chewing of meats, the sight of food, etc., while the parotid gland fails to respond. On the other hand, the latter gland responds with an abundant secretion when dry food, such as dry powdered meat,20 dried bread, etc., is placed in the mouth. With this gland, the inference is that dryness is the active stimulus.
As a digestive secretion, saliva serves several important purposes. By moistening the food it renders mastication and deglutition possible; its natural alkalinity tends to neutralize somewhat such acidity as may be present in the food; it dissolves various solid substances, thus making a solution capable of stimulating the taste nerves; lastly, and most important, it has a marked digestive and solvent action on starchy foods. A large proportion of the non-nitrogenous food consumed by man—in most countries—is composed of some form of starch, and this the body cannot use until it has undergone conversion into soluble forms, such as dextrins and sugar. This it is the function of saliva to accomplish, and it owes its activity in this direction to the presence of a soluble ferment or enzyme known as ptyalin.
Enzymes, which play so important a part in all digestive processes, are a peculiar class of substances produced by the living cells which constitute the various secreting glands. They are of unknown composition, and are peculiar in that the chemical changes they induce are the result of what is termed catalysis, i. e., contact. That is, the enzyme or catalyzer does not enter into the reaction, it is not destroyed or used up, but by its mere presence sets in motion or accelerates a reaction between two other substances. The ordinary illustration from the inorganic world is spongy platinum, which, if placed in contact with a mixture of oxygen and hydrogen, causes the two gases to unite with formation of water, although the two gases alone at ordinary temperature will not so combine. In this reaction the platinum is not altered, neither does it apparently enter into the reaction; it is a simple catalyzer. The chemical nature of the change which most digestive enzymes produce is usually defined as hydrolytic, in which the substance undergoing21 transformation is made to combine with water, thus becoming hydrolyzed, this reaction generally being accompanied by a cleavage or splitting of the molecule into simpler substances. It is to be noted further that enzymes are specific in their action. An enzyme that acts upon starch, for example, cannot act on proteids or fats. Some digestive fluids have the power of producing changes in different classes of foodstuffs, but such diversity of action is always assumed to be due to the presence in the same fluid of different enzymes. Emil Fischer6 has advanced the theory that the specificity of an enzyme is related to the geometrical structure of the substance undergoing change; i. e., that each enzyme is capable of acting upon or attaching itself only to such molecules as have a definite structure with which the enzyme is in harmony. Or, the enzyme may be considered as a key which will fit only into the lock (structure) of the molecule it acts upon.
One characteristic feature of enzymes is the incompleteness of their action. Thus, the enzyme of saliva transforms starch by a series of progressive changes into soluble starch, two or more dextrins, and the sugar maltose as the chief end-product. A mixture of starch paste and saliva under ordinary conditions, however, never results in the formation of a hundred per cent of maltose, but there always remains a variable amount of dextrin which appears to resist further change. This is apparently due to what is known as the reversible action of enzymes. Thus, the chemical reactions involved here are reversible actions, i. e., they take place in opposite directions. The catalyzer not only accelerates or incites a reaction in the direction of breaking down the substance acted upon, but it also aids in the recomposition of the products so formed into the original or kindred substance. With reversible reactions of this sort the opposite changes22 sooner or later strike an equilibrium, which remains constant until some alteration in the conditions brings about an inequality and the reactions proceed until a new equilibrium is established. In the body, however, where the circulating blood and lymph provide facilities for the speedy removal by absorption of the soluble products formed, the reaction may proceed until the original substance undergoing change is completely transformed into the characteristic end-product. This reversible action of enzymes is an important feature, and helps explain certain nutritional changes to be referred to later. Whether all enzymes behave in this way is not as yet determined.
Another peculiarity of digestive enzymes is their extreme sensitiveness to changes in their environment. Powerful in their ability to transform relatively large quantities of a given foodstuff into simple products better adapted for absorption and utilization by the body, they are, however, quickly checked in their action, and even destroyed, when the conditions surrounding them are slightly interfered with. They require for their best action a temperature closely akin to that of the healthy body, and any great deviation therefrom will result at once in an inhibition of their activity. Further, they demand a certain definite reaction of the fluid or mixture, if their working power is to be maintained at the maximum. Indeed, many enzymes, like the ptyalin of saliva, are quickly destroyed if the reaction is greatly changed. Enzymes are thus seen to be more or less unstable substances, endowed with great power as digestive agents, but sensitive to a high degree and working advantageously only under definite conditions. Many perversions of digestion and of nutrition are connected not only with a lack of the proper secretion of some one or more digestive enzyme, but also with the lack of proper surroundings for the manifestation of normal or maximum activity.
23
With these statements before us, we can readily picture for ourselves the initial results following the ingestion of starch-containing foods properly cooked; and it may be mentioned here that the cooking is an essential preliminary, for uncooked starch cannot be utilized in any degree by man. With the mind in a state of pleasurable anticipation, with freedom from care and worry, which are so liable to act as deterrents to free secretion, and with the food in a form which appeals to the eye as well as to the olfactories, its thorough mastication calls forth and prolongs vigorous salivary secretion, with which the food becomes intimately intermingled. Salivary digestion is thus at once incited, and the starch very quickly commences to undergo the characteristic change into soluble products. As mouthful follows mouthful, deglutition alternates with mastication, and the mixture passes into the stomach, where salivary digestion can continue for a limited time only, until the secretion of gastric juice eventually establishes in the stomach-contents a distinct acid reaction, when salivary digestion ceases through destruction of the starch-converting enzyme. Need we comment, in view of the natural brevity of this process, upon the desirability for purely physiological reasons of prolonging within reasonable limits the interval of time the food and saliva are commingled in the mouth cavity? It seems obvious, in view of the relatively large bulk of starch-containing foods consumed daily, that habits of thorough mastication should be fostered, with the purpose of increasing greatly the digestion of starch at the very gateway of the alimentary tract. It is true that in the small intestine there comes later another opportunity for the digestion of starch; but it is unphysiological, as it is undesirable, for various reasons, not to take full advantage of the first opportunity which Nature gives for the preparation of this important foodstuff for future utilization. Further, thorough mastication, by a fine comminution of the food24 particles, is a material aid in the digestion which is to take place in the stomach and intestine. Under normal conditions, therefore, and with proper observance of physiological good sense, a large proportion of the ingested starchy foods can be made ready for speedy absorption and consequent utilization through the agency of salivary digestion.
Nowhere in the body do we find a more forcible illustration of economical method in physiological processes than in the mechanism of gastric secretion. Years ago, it was thought that the flow of gastric juice was due mainly to mechanical stimulation of the gastric glands by contact of the food material with the lining membrane of the stomach. This, however, is not the case, as Pawlow has clearly shown, and it is now understood that the flow of gastric juice is started by impulses which have their origin in the mouth and nostrils; the sensations of eating, the smell, sight, and taste of food serving as psychical stimuli, which call forth a secretion from the stomach glands, just as the same stimuli may induce an outpouring of saliva. These sensations, as Pawlow has ascertained, affect secretory centres in the brain, and impulses are thus started which travel downward to the stomach through the vagus nerves, and as a result gastric juice begins to flow. This process, however, is supplemented by other forms of secretion, likewise reflex, which are incited by substances, ready formed in the food, and by substances—products of digestion—which are manufactured from the food in the stomach. Soups, meat juice, and the extractives of meat, likewise dextrin and kindred products, when present in the stomach, are especially active in provoking secretion. Substances which in themselves have less flavor, as water, milk, etc., are far less effective in this direction, while the white of eggs and bread are entirely without action in directly stimulating secretion. When the latter foods have been in the stomach for a time, however, and the proteid material has25 undergone partial digestion, then absorption of the products so formed calls forth energetic secretion of gastric juice. It is thus seen that there are three distinct ways—all reflex—by which gastric juice is caused to flow into the stomach as a prelude to gastric digestion. Further, it has been shown by Pawlow that there is a relationship between the volume and character of the gastric juice secreted and the amount and composition of the food ingested, thus suggesting a certain adjustment in the direction of physiological economy well worthy of note. A diet of bread, for example, leads to the secretion of a smaller volume of gastric juice than a corresponding weight of meat produces, but the juice secreted under the influence of bread is richer in pepsin and acid, i. e., it has a greater digestive action than the juice produced by meat. The suggestion is that gastric juice assumes different degrees of concentration, with different proportions of acid and pepsin, to meet the varying requirements of a changing dietary.
As has been indicated, pepsin and hydrochloric acid are the important constituents of gastric juice. It is noteworthy, however, that it is the combination of the two that is effective in digestion. Pepsin without acid is of no avail, and acid without pepsin can accomplish little in the digestion of food. Pepsin and acid are secreted by different gland cells in the stomach, and gastric insufficiency, or so-called indigestion, may arise from either a condition of apepsia or from hypoacidity. It is worthy of comment that the amount of hydrochloric acid secreted during 24 hours by the normal individual, under ordinary conditions of diet, amounts to what would constitute a fatal dose of acid if taken at one time in concentrated form. At the outset of gastric secretion, the fluid shows only a slight degree of acidity, but as secretion proceeds, the acidity rises to 0.2–0.3 per cent of hydrochloric acid. The main action of gastric juice is exerted on proteid26 foods, which under its influence are gradually dissolved and converted into soluble products known as proteoses and peptones. It is a process of peptonization, in which the proteid of the food is gradually broken down into so-called hydrolytic cleavage products. The enzyme, like the ptyalin of saliva, is influenced by temperature, maximum digestive action being manifested at about 38° C., the temperature of the body. Further, a certain degree of acidity is essential for procuring the highest degree of efficiency. Ordinarily, it is stated that digestive action proceeds best in the presence of 0.2 per cent hydrochloric acid, but what is more essential for vigorous digestion is a certain relationship between the acid, pepsin, and proteid undergoing digestion. As pepsin and the amount of proteid are increased, the amount of acid, and its percentage somewhat, must be correspondingly increased if digestion is to be maintained at the maximum.
Another important function of gastric juice is that of curdling milk, due to the presence in the secretion of a peculiar enzyme known as rennin. The latter ferment acts upon the casein of milk,—the chief proteid constituent,—transforming it into a related substance commonly called paracasein. This then reacts with the calcium salts present in milk, forming an insoluble curd or calcium compound. From this point on, the digestion of milk-casein by gastric juice is the same as that of any other solid proteid, it being gradually transformed by the pepsin-acid into soluble cleavage products. Why gastric juice should be provided with this special enzyme, capable of acting solely on the casein of milk, can only be conjectured, but we may assume that it has to do with the economical use of this important food. As the sole nutriment of the young, milk occupies a peculiar position as a foodstuff, and being a liquid, its proteid constituent might easily escape complete digestion were it to pass on too hastily through the gastro-intestinal tract. Experiment has shown that when27 liquid food alone is taken into the stomach it is pushed forward into the small intestine in a comparatively short time. Curdled as it is by rennin, however, casein must stay for a longer period in the stomach, like any other solid food, and its partial digestion by gastric juice thereby made certain. For the reasons above stated, it is apparent why milk should not be treated as a drink in our daily diet. Remembering that when milk reaches the stomach it is converted into a solid clot or curd, there is obvious reason for sipping it, instead of taking it by the glassful, thereby favoring the formation of small, individual clots instead of one large curd, and thus facilitating instead of retarding digestion.
Among other factors in gastric digestion, the muscular movements of the stomach walls are to be emphasized, since we have here a mechanical aid to digestion of no small moment, and likewise a means of accomplishing the onward movement of the stomach contents. The outer walls of the stomach are composed of a thick layer of circular muscular fibres, especially conspicuous at the pyloric end of the organ, where the latter is joined on to the intestine; a smaller, less conspicuous layer of longitudinal muscle fibres, and some oblique fibres. At the pylorus, the circular fibres are so arranged as to form a structure which, aided by a peculiar folding of the inner mucous membrane, serves as a sphincter, closing off the stomach from the duodenum, the beginning of the small intestine. The movements of the stomach were first made the subject of careful investigation by Dr. Beaumont in his study of the celebrated case of Alexis St. Martin, a French Canadian, who, in 1822, was accidentally wounded by the discharge of a musket, with the resultant formation of a permanent fistulous opening in the stomach. Dr. Beaumont, in the description7 of his observations, writes that “by28 the alternate contractions and relaxations of these bands (of muscle) a great variety of motion is induced on this organ (the stomach), sometimes transversely, and at other times longitudinally. These alternate contractions and relaxations, when affecting the transverse diameter, produce what are called vermicular or peristaltic motions. . . . When they all act together, the effect is to lessen the cavity of the stomach, and to press upon the contained aliment, if there be any in the stomach. These motions not only produce a constant disturbance, or churning of the contents of this organ, but they compel them, at the same time, to revolve around the interior, from point to point, and from one extremity to the other.” Of more recent investigations, the most important are those made by Cannon,8 with the X-ray apparatus. From these later studies, it is evident that Dr. Beaumont’s view of the entire stomach being involved in a general rotary movement is not correct, since in reality the movements are confined mainly to the pyloric end of the stomach, the fundus or portion nearer the œsophagus not being directly involved. This means that when food material passes into the stomach, it may remain at the fundic end for some time more or less undisturbed before admixture with the gastric juice occurs, and under such conditions, until acidity creeps in, the salivary digestion of starch can continue.
According to the observations of Cannon, the contractile movements of the stomach commence shortly after the entrance of food, the contractions starting from about the middle of the stomach and passing on toward the pylorus. These waves of contraction follow each other very closely, certainly not more than one or two minutes apart, and perhaps less, while the resulting movements bring about an intimate commingling of food and gastric juice in the pyloric29 portion of the stomach; followed by a gradual diffusion of the semi-fluid mixture into the fundus accompanied by a gradual displacement of the more solid food in the latter region. These movements of the stomach are more or less automatic, arising from stimuli—the acid secreted—originating in the stomach itself, although it is considered that the movements are subject to some regulation from extrinsic nerve fibres, such as the vagi and the splanchnics. As digestion proceeds and the mass in the stomach becomes more fluid, the pyloric sphincter relaxes and a certain amount of the fluid material is forced into the intestine by the pressure of the contraction wave. This is repeated at varying intervals, depending presumably in some measure upon the consistency of the mass in the stomach, until after some hours of digestion the stomach is completely emptied.
Especially interesting and suggestive are the experiments made by Cannon9 on the length of time the different types of foodstuffs remain in the stomach. Using cats as subjects, he found that fats remain for a long period in the stomach; they leave that organ slowly, the discharge into the intestine being at about the same rate as the absorption of fat from the small intestine or its passage into the large intestine. Carbohydrate foods, on the other hand, begin to leave the stomach soon after their ingestion. They pass out rapidly, and at the end of two hours reach a maximum amount in the small intestine almost twice the maximum for proteids, and two and a half times the maximum for fats, both of which maxima are reached only at the end of four hours. Carbohydrates remain in the stomach about half as long as proteids. Proteids, Cannon finds, frequently do not leave the stomach at all during the first half-hour after they are eaten. After30 two hours, they accumulate in the small intestine to a degree only slightly greater than that reached by carbohydrates an hour and a half earlier. The departure of proteids from the stomach is therefore slower at first than that of either fats or carbohydrates. When a mixture of equal parts of carbohydrates and proteids is fed, the discharge from the stomach is intermediate in rapidity. When fat is added to either carbohydrates or proteids it retards the passage of both foodstuffs through the pylorus.
It is evident from what has been stated that the gastric digestion of proteid foods is a comparatively slow process, involving several hours of time; and further, that food material in general remains in the stomach for varying periods, dependent upon its chemical composition. It would appear further, that relaxation of the pyloric sphincter, allowing passage of chyme into the intestine, must depend somewhat upon chemical stimulation, as this offers the most plausible explanation of the diversity of action seen with the different foodstuffs. As has been pointed out, gastric digestion is primarily a process for the conversion of proteid food into soluble products. It would be a mistake, however, to assume that the digestion of proteid foods is complete in the stomach. Stomach digestion is to be considered more as a preliminary step, paving the way for further changes to be carried forward by the combined action of intestinal and pancreatic juice in the small intestine. The importance of gastric digestion is frequently overrated. It is unquestionably an important process, but not absolutely essential for the maintenance of life. Dogs have lived and flourished with their stomachs removed, the intestine being joined to the œsophagus. The intestine is a much more important part of the alimentary tract; it is likewise far more sensitive to changing conditions than the stomach, and undoubtedly one function of the latter organ is to protect the intestine and preserve it from insult.31 The stomach may be compared to a vestibule or reservoir, capable of receiving without detriment moderately large amounts of food, together with fluid, in different forms and combinations, with the power to hold them there until by action of the gastric juice they are so transformed that their onward passage into the intestine can be permitted with perfect safety. Then, small portions of the properly prepared material may be discharged from time to time through the pylorus without danger of overloading the intestine, and in a form capable of undergoing rapid and complete digestion. Further, the stomach as a reservoir is very useful in bringing everything to a proper and constant temperature before allowing its entry into the intestine. Another fact of some importance is that, contrary to the general view, absorption from the stomach of the products of digestion is not very rapid under ordinary conditions. Even water and soluble salts pass very slowly into the circulation from the stomach. Like the partially digested food material, they are carried forward through the pyloric sphincter into the intestine, where absorption of all classes of material is most marked.
It is in the small intestine that both digestion and absorption are seen at their best. It is here that all three classes of foodstuffs are acted upon simultaneously through the agency of the pancreatic juice, intestinal juice, and bile. Here, too, are witnessed some of the most complicated and interesting reactions and changes occurring in the whole range of digestive functions. Especially noteworthy is the peculiar mechanism by which the secretion of pancreatic juice is set up and maintained. On demand, pancreatic juice is manufactured in the pancreas and poured into the intestine just beyond the pylorus through a small duct—the duct of Wirsung. Secretion is started by contact of the acid contents of the stomach with the mucous membrane of the small intestine, so that as soon as the acid chyme passes through the pyloric sphincter32 there commences an outflow of pancreatic juice into the intestine. While acid is plainly the inciting agent in this secretory process, its action is indirect. It does not cause secretion through reflex action on nerve fibres, but it acts upon a substance formed in the mucous membrane of the intestine, transforming it into secretin, which is absorbed by the blood and carried to the pancreas, where it excites secretory activity. As would be expected from the foregoing statements, the secretion of pancreatic juice commences very soon after food finds its way into the stomach, and naturally increases in amount with the onward passage of acid chyme into the intestine, the maximum flow being obtained in the neighborhood of the third or fourth hour, after which the secretion gradually decreases. In man, it is estimated on the basis of one or two observations that the amount secreted during 24 hours is about 700 cc., or a pint and a half. Careful experiments, however, tend to show that the quantity of secretion depends in some measure at least upon the character of the food, and also that the composition of the secretion varies with the character of the food. Thus, on a diet composed mainly of meat, the proteid-digesting enzyme is especially conspicuous, while on a bread diet, with its large content of starch, the starch-digesting enzyme is increased in amount. In other words, there is suggested the possibility of an adaptation in the composition of the secretion to the character of the food to be digested.
Pancreatic juice is an alkaline fluid, rather strongly alkaline in fact, from its content of sodium carbonate, and is especially characterized by the presence of at least three distinct enzymes; viz., trypsin, a proteid-digesting ferment; lipase, a fat-splitting enzyme; and amylopsin, a starch-digesting enzyme. It has already been pointed out how dependent the secretion of pancreatic juice is upon the co-operation of the intestinal mucous membrane. A similar33 dependence is found when the digestive activity of the secretion is studied. As just stated, pancreatic juice contains a proteid-digesting enzyme. This statement, however, is not strictly correct, for if the secretion is collected through a cannula so that it does not come in contact with the mucous membrane of the intestine, it is found free from any digestive action on proteids. The secretion is activated, however, by contact with the duodenal membrane. Expressed in different language, pancreatic juice as it is secreted by the gland does not contain ready-formed trypsin; it does contain, however, an inactive pro-enzyme, which, under the influence of a specific substance contained in the intestinal mucous membrane, known as enterokinase, is transformed into the active enzyme trypsin. There is thus seen another suggestive example of the close physiological relationship between the small intestine and the activity of the pancreatic gland, or its secretion.
The chemical changes taking place in the small intestine are many and varied. The acid chyme, with its admixture of semi-digested food material, as it passes through the pyloric sphincter into the small intestine, is at once brought into immediate contact with bile, pancreatic juice, and intestinal juice, all of which are more or less alkaline in reaction. As a result, the acidity of the gastric juice is rapidly overcome, and the enzyme pepsin, which up to this point could exert its characteristic digestive action, is quickly destroyed by the accumulating alkaline salts. Pepsin digestion thus gives way to trypsin digestion,—most effective in an alkaline medium,—and the proteids of the food, already semi-digested by pepsin-acid, are further transformed by trypsin; aided and abetted by another enzyme, known as erepsin, secreted by the mucous membrane of the intestine. These two enzymes are much more powerful agents than pepsin. It is true that they begin work where pepsin left off, but most striking is34 the character of the end-products which result from their combined action, since they are small molecules and there is a surprising diversity of them. In other words, while gastric digestion breaks down the proteid foodstuffs into soluble bodies, such as proteoses and peptones closely related to the original proteids, in pancreatic digestion as it takes place in the intestine there is a profound breaking down, or disruption of the proteid molecule into a row of comparatively simple nitrogenous fragments, many of them crystalline bodies; such as leucin, tyrosin, glutaminic acid, aspartic acid, arginin, lysin, histidin, etc., known chemically as monoamino-acids and diamino-acids. We have no means of knowing to how great an extent these more profound disruptive changes of the proteid molecule take place in the intestine. Whether practically all of the ingested proteid food is broken down into these relatively simple compounds prior to absorption, or whether only a small fraction suffers this change, cannot be definitely stated.
A few years ago, the majority of physiologists held to the view that in the digestion of proteid food all that was essential was its conversion into soluble and diffusible forms which would permit of ready absorption into the blood. The belief was prevalent that, since the proteid of the food was destined to make good the proteid of the blood and through the latter the proteids of the tissues, any change beyond what was really necessary for absorption of the proteid would be uneconomical and indeed wasteful. On the other hand, due weight must be given to the fact that in trypsin digestion, proteid can be quickly broken down into simple nitrogenous compounds, and that in the enzyme erepsin, present in the mucous membrane of the intestine, we have an additional ferment very efficient in bringing about cleavage of proteoses and peptone into amino-acids. From these latter facts it might be argued that, in the digestion of proteid foodstuffs by the35 combined action of gastric and pancreatic juice in the alimentary tract, a large proportion of the proteid is destined to undergo complete conversion into amino-acids, and that from these fragments the body, by a process of synthesis, can construct its own peculiar type of proteid.
This latter suggestion is worthy of a moment’s further consideration. As is well known, every species of animal has its own particular type of proteid, adapted to its particular needs. The proteids of one species directly injected into the blood of another species are incapable of serving as nutriment to the body, and frequently act as poisons. Man in his wide choice of food consumes a great variety of proteids, all different in some degree from the proteids of his own tissues. Is it not possible, therefore, that it is the true function of pancreatic and intestinal digestion to break down the different proteids of the food completely into simple fragments, so that the body can reconstruct after its own particular pattern the proteids essential for its nourishment? Or, we can follow the suggestion contained in the work of Abderhalden,10 who finds that in the long continued digestion of various proteids by pancreatic juice there results in addition to the amino-acids a very resistant residue, non-proteid in nature, which is termed polypeptid. In other words, Abderhalden believes that pepsin, trypsin, and erepsin are not capable of bringing about a complete breaking down of proteids into amino-acids, but that there always remains a nucleus of the proteid not strictly proteid in nature, though related thereto,—polypeptid,—which may serve as a starting-point for the synthesis or construction of new proteid molecules, the various amino-acids being employed to finish out the structure and give the particular character desired. This view, however, is rendered somewhat untenable by the more recent experiments36 of Cohnheim,11 who claims that proteids can be completely broken down by pepsin, trypsin, and erepsin, and consequently polypeptids would hardly be available for the synthesis of proteids. Moreover, Bergell and Lewin12 have ascertained that there is present in the liver an enzyme or ferment which has the power of digesting or breaking down certain dipeptids and polypeptids into amino-acids. Hence, it follows that if any polypeptids are absorbed from the intestine, they would naturally be carried to the liver, where further cleavage into fragments suitable for synthetical processes might occur. In any event, there is good ground for the belief that the more or less complete disruption of the proteid molecule into small fragments renders possible a synthetical construction of new proteid to meet the demands of the organism; a fact of great importance in our conception of the possibilities connected with this phase of proteid nutrition.
Fatty foods undergo little or no chemical alteration until they reach the small intestine. During their stay in the stomach they naturally become liquid from the heat of the body, and there is more or less liberation of fat from the digestive action of gastric juice on cell walls, connective tissues, etc. Most food fat is in the form of so-called neutral fat, which must undergo hydrolysis or saponification before it can be absorbed and thus made available for the body. This is accomplished by the enzyme lipase, or steapsin, of the pancreatic juice, aided indirectly by the presence of bile. Under the influence of this fat-splitting enzyme all neutral fats, whether animal or vegetable, are broken apart, through hydrolysis, into glycerin and a free fatty acid; the latter reacting in some measure with the sodium carbonate of37 the pancreatic juice to form a sodium salt, or soluble soap, while perhaps the larger part of the fatty acid is held in solution by the bile present. Soap, free acid, and glycerin are then absorbed from the intestine and are found again combined in the lymph as neutral fat. In this way the fats of the food are rendered available for the nourishment of the body.
The next important chemical change taking place in the small intestine is that induced by the amylopsin of the pancreatic juice, which, acting in essentially the same manner as the ptyalin of saliva, converts any unaltered starch into dextrins and sugar. The latter substance, maltose, is exposed to the action of another enzyme contained in the intestinal secretion termed maltase, which transforms it into dextrose, a monosaccharide.
In these ways the proteids, fats, and carbohydrates of the food are gradually digested, so far as conditions will admit, digestion being practically completed by the time the material reaches the ileocæcal valve at the beginning of the large intestine. Throughout the length of the small intestine absorption proceeds rapidly; water, salts, and the products of digestion passing out from the intestine into the circulating blood and lymph. At the ileocæcal valve, however, the contents of the intestine are practically as fluid as at the beginning of the small intestine, due to the fact that water is continually being secreted into the intestine. In the large intestine, the contents become less and less fluid through reabsorption of the water, and as the propulsive movements of the circular and longitudinal muscle fibres of the intestinal wall carry the material onward toward the rectum, the last portions of available nutriment are absorbed. Finally, in varying degree, certain putrefactive changes are observed in the large intestine involving a breaking down of some residual proteid matter, through the agency of micro-organisms almost invariably present, with formation of such substances38 as indol, skatol, phenol, fatty acids, etc. These processes, however, in health are held rigidly in check, and count for little in fitting the food for absorption. Digestion, on the other hand, extending as we have seen from the mouth cavity to the ileocæcal valve, is the handmaiden of nutrition, preparing all three classes of organic foodstuffs for their passage into the circulating blood and lymph, and thus paving the way for their utilization by the hungry tissue cells.
39
ABSORPTION, ASSIMILATION, AND THE PROCESSES OF METABOLISM
Topics: Physiological peculiarities in absorption. Chemical changes in epithelial walls of intestine. Two pathways for absorbed material. Function of the liver as a regulator of carbohydrate. Absorption of proteid products. Assimilation of food products. Anabolism. Katabolism. Metabolism. Processes of metabolism. Older views regarding oxidation. Discoveries of Lavoisier. The views of Liebig. Theory of luxus consumption. Oxidation in the body not simple combustion. Oxygen not the cause of the decompositions. Oxidation not confined to any one place. Intracellular enzymes. Living cells the guiding power in katabolism. Some intermediary products of tissue metabolism. Chemical structure of different proteids. Decomposition products of nucleoproteids. Relation to uric acid. Action of specific intracellular enzymes. Creatin and creatinin. Relation to urea. Proteid katabolism a series of progressive chemical decompositions. Intracellular enzymes as the active agents.
Digestion being completed, and the available portion of the foodstuffs thereby converted into forms suitable for absorption, the question naturally arises, In what manner are these products transported from the alimentary tract to the tissues and organs of the body? In attempting to answer this question, we shall find many illustrations of the precise and undeviating methods which prevail in the processes of nutrition. For example, it would seem plausible to assume that the different forms of sugar entering into man’s ordinary diet, all of them being soluble, would be directly absorbed and at once utilized, but such is far from being the case. Milk-sugar and cane-sugar, both appearing in greater40 or less degree in our daily dietaries, if introduced directly into the blood, are at once excreted through the kidneys unchanged. The body cannot use them, and they are gotten rid of as speedily as possible, much as if they were poisons. When taken by way of the mouth, however, they are utilized, simply because in the intestine two enzymes are present there, known as lactase and invertase, which break each of the sugars apart into two smaller molecules. In other words, milk-sugar and cane-sugar are disaccharides, and if they are to be absorbed in forms capable of being made use of by the body they must be split apart into simpler sugars, viz., monosaccharides, such as dextrose, levulose, etc. The great bulk of the carbohydrate food consumed by man is in the form of starch, and this, as we have seen, is converted into maltose by the action of saliva and pancreatic juice. Maltose, however, like cane-sugar, is a disaccharide, and the body has no power to burn it or utilize it directly; but in the intestine and elsewhere is an enzyme termed maltase, which breaks up maltose into two molecules of the monosaccharide dextrose, and this the body can use. Man frequently consumes starch to the extent of a pound a day, and if utilized it must all undergo transformation into maltose, and then into dextrose. There is no apparent reason why maltose should not be absorbed and assimilated as readily as dextrose, but so urgent is the necessity for this conversion into dextrose that in the blood itself there is present maltase, to effect the transformation of any maltose that may gain entrance there. We are here face to face with a simple fact in nutrition. The body cannot utilize disaccharides directly. Why it is so we cannot say, but the fact is a good illustration of the principle that nothing can be taken for granted in our study of nutrition.
For years, physiologists assumed that the ordinary physical laws of osmosis, imbibition, and diffusion were quite adequate41 to explain the passage of digested food materials into the blood and lymph. If a substance was soluble and diffusible, that was sufficient; it would quite naturally be absorbed in harmony with its diffusion velocity. This, however, is not wholly true, since experiment shows that the rapidity of absorption of diffusible substances through the wall of the intestine is by no means always proportional to the diffusion velocity of the substance. The lining membrane of the small intestine, where absorption mainly takes place, is not to be compared to a dead parchment membrane. On the contrary, it is made up of living protoplasmic cells; absorption is not a physical, but a physiological, process, in which the living epithelium cells stand as guardians of the portals, ready to challenge and, if need be, modify the rate of passage. Osmosis and diffusion undoubtedly play some part in absorption, but they alone are not sufficient to account for what actually takes place in the absorption of digestion products, and other substances from the living intestine.
The primary products formed in the digestion of proteid foods—the proteoses and peptones—afford another illustration of physiological peculiarity in absorption. These bodies are readily soluble and quite diffusible, yet they are never found to any extent in the circulating blood and lymph during health. It is of course possible, as has been previously suggested, that as soon as formed they undergo transformation into simpler decomposition products in the small intestine; but this is by no means certain. If proteoses and peptones are injected directly into the blood, they cause a marked disturbance, influencing at once blood-pressure, affecting the coagulability of the blood, and in many other ways exhibiting a pronounced deleterious action which at once indicates they are out of their normal environment. They are not at home in the circulating blood, and the latter medium gets rid of them as speedily as possible; they behave like42 veritable poisons, and yet they are the primary products formed in the digestion of all proteid foodstuffs. On the basis of all physical laws governing diffusion they should be absorbed, and help to renew the proteids of the blood and later the proteids of the tissues. Yet, as we have said, they are not normally present in the blood or lymph. Apparently, in the very act of absorption, as they pass through the epithelial cells of the intestinal wall, before they gain entrance to the blood stream, they undergo transformation into serum-albumin and globulin, the characteristic blood proteids. The other alternative is that, as previously mentioned, they are completely broken down in the intestine into amino-acids, etc., and these simpler products synthesized, as they pass through the intestinal wall toward the blood, into serum-albumin and globulin. Certainly as yet, there is no evidence that the amino-acids, as such, go through the epithelial cells of the intestine; they are not found in the blood or lymph to any appreciable extent, yet the proteids of the blood are reinforced in some manner by the products of proteid digestion. Whichever view is correct, one thing is perfectly obvious, viz., that in the act of absorption the products resulting from the gastric and pancreatic digestion of proteid foods are exposed to some influence, presumably in the epithelial cells of the intestinal wall, by which there is a reconstruction of proteid. Further, the proteid substances so formed are of the type peculiar to the blood of that particular species of animal. The proteids of beef, mutton, chicken, oatmeal, or bread go to make the proteids of human blood.
From these statements, it is obvious that what we term absorption is something more than a simple diffusion of soluble substances from the alimentary tract into the blood current. The process is much more complex than appears on the surface, and our lack of definite knowledge, in spite of numerous efforts to unravel the mystery, merely strengthens the view43 that we are dealing here with an obscure physiological problem, and not a simple physical one. Digestion induces a splitting up of the food proteid into fragments, large or small, while incidental to absorption there is apparently a reconstruction, or synthesis, of proteid from the fragments so formed. The process seems somewhat costly, physiologically speaking, yet when one considers the variety of proteids consumed as food, it is easy to comprehend how essential it is that in some manner, as in absorption, there be opportunity for construction of the specific proteids of the blood and lymph.
We find an analogous process in the absorption of fats. As we have seen, the fats of the food are broken apart in the small intestine into glycerin and free fatty acid, a portion of the latter, and perhaps all, combining with the alkali of the intestinal juices to form soluble soaps, or sodium salts of the respective fatty acids. The neutral fats present in animal and vegetable foods are all alike in containing the glyceryl radicle, but they differ in the character of the fatty acids present. Further, one form of animal fat, like that from beef, may contain quite a different proportion of stearin, palmitin, and olein than is present in the fat of another animal, like mutton. By digestion, however, they are all broken apart into fatty acid and glycerin. These acids and their salts can be readily detected in the intestine, but they are not found in the blood or lymph, yet shortly after fatty food is taken the lymph is seen to be milky from fat. Obviously, the fatty acids liberated in the intestine are absorbed, either as soluble soaps or as free fatty acids dissolved in bile, but as they pass through the epithelial cells of the intestine into the lacteal radicles, there is a synthesis or reconstruction of fat; and as a result, neutral fats and not soaps are found in the lymph. Here, then, we have a process quite analogous to what apparently occurs in the absorption of proteid, though less complex; and it is possible that this is one of the factors which44 aids in the formation of a specific fat mixture corresponding, in a measure, to the type of fat present in the particular species. It is well understood that the fat of an animal’s tissues may be modified somewhat by the character of the fat fed, yet in spite of this there is a certain degree of constancy in composition which calls for explanation. Sheep and oxen feeding in the same pasture have fat widely different in the proportion of stearin, palmitin, etc. The fat of man’s tissues is fairly definite in composition, yet he eats a great variety of fatty foods. One man may consume large amounts of hard mutton fat with its relatively large content of stearin, while another individual may take his fat mainly in the form of the soft butter fats, with their relatively large content of olein and palmitin. In both cases, the fat of the man’s tissues will be essentially the same. To be sure, the changes that take place in the tissue cells, reinforced by the construction of fat from other sources, may be partly responsible for this constancy of composition, but the transformations incidental to absorption are quite possibly, in some measure, helpful thereto.
The great bulk of the digested food material is absorbed from the small intestine, and there are two pathways open through which the absorbed material can gain access to the blood. The one path leads directly to the liver, and substances taking this course are exposed to the action of this organ, before they enter into the general circulation. The other path is through the lacteal or lymphatic system, and constitutes a roundabout way for substances to enter the blood stream, since they must first pass through the thoracic duct before entering the main circulation. As a general truth, it may be stated that fats are absorbed through the latter channel, while carbohydrates and proteids follow the first path. The innumerable blood capillaries in the villi of the intestine take up the products resulting from the digestion45 of proteids and carbohydrates, through which they are passed into the portal vein, and thereby distributed throughout the liver. This means that both carbohydrates and proteids—or their decomposition products—are exposed to a variety of possible changes in this large glandular organ, before they can enter into the tissues of the body. As we have seen, practically all carbohydrate food is converted into a monosaccharide, principally dextrose, in the alimentary tract; and it is in this form of a simple sugar that the carbohydrate passes into the blood. This might easily mean a pound of sugar absorbed during the twenty-four hours, and would obviously give to the blood a high degree of concentration, unless the excess was quickly disposed of. Sugar is very diffusible, and if it accumulates to any extent in the blood it is quickly gotten rid of by excretion through the kidneys. This, however, is wasteful, physiologically and otherwise, and does not ordinarily occur except in diseased conditions. Further, physiologists have learned that a certain small, but definite, amount of sugar in the blood is a necessary requirement in nutrition, and it is the function of the liver to maintain the proper carbohydrate level.
We must again emphasize the great importance of carbohydrate food; there is a far larger amount of starchy food consumed than of any other foodstuff, and it is more readily available as a source of energy. Its presence in the blood, in the form of sugar, is constantly demanded, but it must be kept within the proper limits for the uses of the different tissues and organs of the body. The liver serves as an effective regulator, maintaining, in spite of all fluctuations in the supply and demand, a definite percentage of sugar such as is best adapted to keep the tissues of the body in a normal and healthy condition. This regulation by the liver is rendered possible through the ability of the hepatic cells to transform the sugar brought to the gland into glycogen, so-called animal46 starch, which is stored up in the liver until such time as it is needed by the body. The process is one of dehydration, the reverse of what takes place in the intestine when ordinary starch is converted into maltose and dextrose. The efficiency of this regulating mechanism depends also upon the ability of the liver to transform glycogen into sugar, presumably through the agency of an enzyme in the hepatic cells. Hence, glycogen may be looked upon as a temporary reserve supply of carbohydrate, manufactured and stored in the liver during digestion, when naturally large amounts of sugar are passing into the portal blood, and to be drawn upon whenever from any cause the content of sugar in the blood threatens to fall below normal. Obviously, there must be some delicate machinery for the adjustment of these opposite changes in the liver, and we may well believe that it is associated with the composition of the blood itself, which in some fashion stimulates and inhibits, as may be required, the functional activity of the liver, or its component cells. In any event, we have in this so-called glycogenic function of the liver a most effective means for accomplishing the complete and judicious utilization of all the sugar formed from the carbohydrates of the food, after it has once passed beyond the confines of the alimentary tract into the blood; preventing all loss, and at the same time guarding against all danger, from undue accumulation of sugar in the circulation. We see, too, how wise the provision that all sugar should pass from the alimentary canal into the portal circulation and not by way of the lymphatics, since by the latter channel the regulating action of the liver would be mainly lost. Further, recalling how soluble and diffusible sugar is, we may well marvel that it practically all passes from the intestine by way of the blood, and escapes entry into the lymphatics. Surely, this marked shunning of the other equally accessible pathway affords a striking illustration of selective action such as47 might be expected in a physiological process, but not in harmony with the ordinary physical laws of osmosis or diffusion. In conformity with this statement, it may be mentioned that appropriate experiments have clearly demonstrated that the different sugars available as food are not absorbed from the intestine in harmony with their diffusion velocity, but show deviations therefrom which can be explained only on the ground that the intestinal wall exercises some selective action, due to the living cells composing it. Likewise interesting in their bearing on nutrition are the observations of Hofmeister,13 who finds by experiments on dogs that the assimilation limit of the different sugars shows marked variation. Thus, dextrose, levulose, and cane-sugar have the highest assimilation, while milk-sugar is far less easily and completely assimilated. If this is equally true of man, it indicates that starchy foods, with their ultimate conversion into dextrose, are to be ranked as having a high assimilation limit, thus affording additional evidence of their high nutritive value.
In the absorption of proteid products, their passage from the intestine by way of the portal circulation insures exposure to the action of the hepatic cells, before they are distributed by the general circulation throughout the body. It is only under conditions of an excessive intake of proteid foods that their products are absorbed by way of the lymphatics. These points are clearly established, and there is every ground for believing that substantial reasons exist to account for this single line of departure. Just what the liver does, however, is uncertain. In fact, as already indicated, there is lack of definite knowledge as to how far the proteid foods are broken down in digestion, prior to absorption. The combined action of pepsin, trypsin, and erepsin, if sufficiently long continued, can accomplish a complete disruption of the proteid molecule.48 We are inclined to assume in a general way that the “proteids taken as food cannot find a place in the economy of the animal body till they have been, as it were, melted down and recast.”14 How far this melting down or disruption extends in normal digestion, we do not at present know. As already stated, neither proteoses and peptones, nor the amino-acids, are found in the blood stream in sufficient amounts, or with that frequency, to suggest absorption in these forms. Possibly, as some physiologists have suggested, the amount of any of these products to be found at any one time in a given quantity of blood is too small for certain recognition, yet in the twenty-four hours the amount passing from intestine to liver might be sufficiently large to equal the total proteid absorbed. We can, however, at present only conjecture, and must rest content with the simple statement that in the digestion of the proteid foodstuffs, proteoses, peptones, and amino-acids are formed, and that by transformation or total reconstruction of these products, special types of proteid are manufactured either in the epithelial cells of the intestinal walls during absorption, or elsewhere in the body after absorption. If this latter is the case, the liver might readily be regarded as a likely spot for the synthesis to occur.
Bearing in mind what has been said regarding the production of specific types of proteid by every species of animal, we can the more readily conceive of a synthesis “out of fragments of the original molecules rearranged and put together in new combinations, by processes in which the intestine can hardly be supposed to play a part.” This, the liver might well be assumed as capable of accomplishing, and if we were disposed to accept this view we might use as an argument the fact that the products of proteid digestion are taken directly to this organ, before being cast loose in49 the tissues and organs of the body. There is perhaps as good ground for assuming that a synthesis or reconstruction of proteid takes place all over the body; that, as suggested by Leathes, “the synthesis of proteids is a function of every cell in the body, each one for itself, and that the material out of which all proteids in the body are made is not proteid in any form, but the fragments derived from proteids by hydrolysis, probably the amido-acids, which in different combinations and different proportions are found in all proteids, and into which they are all resolved by the processes, autolytic or digestive, which can be carried out in every cell in the body.” It is certainly a reasonable hypothesis, and since we lack positive knowledge it cannot at present be disproved. All that we can affirm in the light of established fact is that the products of proteid digestion are absorbed from the intestine by way of the portal circulation, and that either in their passage through the intestinal wall, or later on in the liver or elsewhere, there is a construction of new proteid to meet the wants of the body. The liver, indeed, may be effective in both construction and destruction of proteid, but there is no way of telling at present just how far it acts in either direction.
Regarding the absorption of fats, a single statement will suffice, in addition to what has already been said. Fats gain access to the general circulation by passing from the intestine into the lacteal radicles, thence into the lymphatics, whence they move onward into the thoracic duct, and from there are emptied into the great veins at the neck. A small amount is apparently absorbed in the form of soap by the portal circulation, but by far the larger amount of fat gains access to the blood stream without going through the liver.
In these ways, the blood and lymph are continually supplied with proteid, fat, and carbohydrate from the ingested50 food, and as these fluids surround and permeate the organized elements of the tissues, the latter are enabled to gain what they need to maintain their nutritive balance. Living matter is essentially unstable; it is the seat of chemical changes of various kinds, anabolic or constructive, and katabolic or destructive. The more comprehensive term “metabolic” is applied to all of these changes that take place in living matter. In anabolism, the dead, inert proteids, fats, and carbohydrates are more or less assimilated and made a part of the living matter of the tissue cells, while at the same time a certain amount of the food material, probably the larger amount, is simply stored as such, or left to circulate in the blood and lymph, without being raised to the higher level of living protoplasm. In katabolism, this accumulated material, and in some degree the living substance itself, is broken down or disintegrated with liberation of the stored-up energy, which manifests itself in the form of heat and mechanical work. At times, the anabolic processes predominate and there is a relatively large accumulation of stored-up materials; while at other times, katabolism, with its attendant chemical decompositions, predominates, and the body loses correspondingly. The point to be emphasized here is that the living body, with its multitude of living cells, is the seat of incessant change. Construction and destruction are continually going forward side by side; sometimes the one and sometimes the other predominating, according to existing conditions. The living protoplasm with its attendant storage material is, under ordinary conditions, constantly being made good from the assimilated food, a part of which is raised to the dignity of living matter and becomes an integral part of the living cells, while the larger portion is simply stored for future uses, or circulates in the blood and lymph which bathe them. Doubtless, this storage or circulating material is the main source of the energy which constantly flows from the cells51 in the form of heat and of work, as a result of the disruptive changes that constitute katabolism.
Worthy of special notice is the fact that cell protoplasm is essentially proteid in nature; water and proteid make up the larger part of its substance, to which are added small proportions of carbohydrate, fat, and mineral matter. Proteid is the basis of cell protoplasm; it is the chemical nucleus of living matter, and owing to the large size of its molecule, with its large number of contained atoms, is capable of many combinations and many alterations. Most of the reactions characteristic of katabolism centre around this proteid, but the disruptive changes that occur undoubtedly involve more largely the circulating materials present in the blood and lymph, and which bathe the cells, rather than the so-called fixed, or organ proteid, of the cell substance itself. Still, while the circulating blood and lymph furnish largely the substances which are made to undergo disintegration in katabolism, the living protoplasmic cell is the controlling power which regulates the extent and character of the decompositions, and proteid matter is the chemical basis of protoplasm. From these statements, we again have suggested the significant importance of the proteid foods in nutrition, since they alone can furnish the material which constitutes the chemical basis of living cells. The human body, which represents the highest form of animal life, is merely, as stated by another, “literally a nation of cells derived from a single cell called the ovum, living together, but dividing the work, transformed variously into tissues and organs, and variously surrounded by protoplasm products” (Waller).
The processes involved in metabolism are not easily unravelled. The word itself is simple, but it is employed to designate that complex of “chemical changes in living organisms which constitute their life, the changes by which their52 food is assimilated and becomes part of them, the changes which it undergoes while it shares their life, and finally those by which it is returned to the condition of inanimate matter. Gathered together under this one phrase are some of the most intricate and inaccessible of natural phenomena. It implies also, and gently insists on the idea, that all the phenomena of life are at bottom chemical reactions” (Leathes). Regarding the processes of anabolism, as in the construction of living protoplasm out of inert food materials, we can say nothing. This is altogether beyond our ken at present, and doubtless will remain so, since it involves a chemical alteration, or change, akin to that of bringing the dead to life. With the processes of katabolism, however, we may hope for more satisfactory results; and, indeed, to-day we have considerable information of value as to some of the methods, at least, which are the cause of this phase of nutrition. This knowledge, however, has been slow of attainment.
In the earlier years of the sixteenth century, when anatomy and physiology were beginning to make progress, the savants of that day, hampered as they were by grave misconceptions and by the lack of any understanding of chemical phenomena, could not take advantage, naturally, of the suggestion that as wood burns or oxidizes in the air with liberation of heat, so might the food substances, absorbed by the body, undergo oxidation in the tissues and thus give rise to animal heat. Such suggestions were at that time as a closed book, and so we find Vesalius, in 1543, teaching the Galenic doctrines in physiology then prevalent. The conception of heat production, as it existed at that time, may be inferred from the following quotation:15 “The parts of the food absorbed from the alimentary canal are carried by the portal blood to the53 liver, and by the influence of that great organ are converted into blood. The blood thus enriched by the food is by the same great organ endued with the nutritive properties summed up in the phrase ‘natural spirits.’ But blood thus endowed with natural spirits is still crude blood, unfitted for the higher purposes of the blood in the body. Carried from the liver by the vena cava to the right side of the heart, some of it passes from the right ventricle through innumerable invisible pores in the septum to the left ventricle. As the heart expands it draws from the lungs through the vein-like artery air into the left ventricle. And in that left cavity, the blood which has come through the septum is mixed with the air thus drawn in, and by the help of that heat, which is innate in the heart, which was placed there as the source of the heat of the body by God in the beginning of life, and which remains there until death, is imbued with further qualities, is laden with ‘vital spirits,’ and so fitted for its higher duties. The air thus drawn into the left heart by the pulmonary vein, at the same time tempers the innate heat of the heart and prevents it from becoming excessive.” In other words, heat was considered as a divine gift, and as can readily be seen, there was an utter lack of appreciation of the use of air in breathing. Even van Helmont, who lived in 1577–1644, and was in a sense an alchemist, still gave credence to the spirits, viz., that the food absorbed from the stomach and intestine is in the liver endued with natural spirits, while in the heart the natural spirits are converted into vital spirits, and in the brain the vital spirits are transformed into animal spirits.16 Later, Malpighi discovered the true structure of the lungs, and Borelli, in 1680, exposed the erroneous views then prevalent regarding the purpose of breathing. It is not true, says Borelli, that the use of breathing is to cool the excessive54 heat of the heart or to ventilate the vital flame, but we must believe that this great machinery of the lungs, with their accompanying blood vessels, is for some grand purpose. In a long and vigorous argument, he contends that the “air taken in by breathing is the chief cause of the life of animals, far more essential than the working of the heart and the circulation of the blood.” He quotes the experiments of Boyle, who showed in 1660 “that even in a partial vacuum brought about by his air pump, flame was extinguished and life soon came to an end; the candle went out and the mouse or the sparrow died.”
At this time, and for long afterwards, the belief was prevalent that the air taken up by the blood in the lungs was the air of the atmosphere in its entirety. No one appears to have thought of the possibility of only a part of the air being used, for at that time there was no suspicion that air was a mixture of substances. Mayow, however, in 1668, showed that it was not the whole air which was employed for respiration, but a particular part only. At this time, great attention was being given to a study of nitre or saltpetre; its wonderful properties in combustion were being recognized, and Mayow, who was a chemist of repute, claimed that it had its origin partly in the air and partly in the earth. The air “which surrounds us, and which, since by its tenuity escapes the sharpness of our eyes, seems to those who think about it to be an empty space, is impregnated with a certain universal salt, of a nitro-saline nature, that is to say, with a vital, fiery, and in the highest degree fermentative spirit,” to which the name of “igneo-aereus” was applied. Nitre was shown to be composed of a sal fixum or sal alkali,—potash as it is now called,—and was obviously derived from the earth, while the other part of nitre was made up of the spiritus acidus, or nitric acid. For a time it was supposed that the whole of this spiritus acidus was contained in the atmosphere,55 but it was soon recognized that this could not be the case, since nitric acid was found to be a corrosive liquid, destructive to life and quite incapable of supporting combustion. Hence, Mayow concluded that only a part of the acid exists in the atmosphere, viz., that part which he termed spiritus nitro-aereus. In combustion, there is something in the air which is necessary for the burning of every flame, unless perchance igneo-aereal particles should pre-exist in the thing to be burnt. These igneo-aereal particles form “the more active and subtle part of air which is thus necessary for combustion, exist in nitre and indeed constitute its ‘more active and fiery part.’” Mayow fully recognized that burning and breathing involved in a measure the same process; both consisted in the consumption of the igneo-aereal particles present in the air. “If a small animal and a lighted candle be shut up in the same vessel, the entrance into which of air from without be prevented, you will see in a short time the candle go out, nor will the animal long survive its funeral torch. Indeed, [says Mayow] I have found by observation that an animal shut up in a flask together with a candle will continue to breathe for not much more than half the time than it otherwise would, that is, without the candle.” Something contained in the air, necessary alike for supporting combustion and for sustaining life, passes from the air into the blood. Mayow expressed his thoughts in these words: “And indeed it is very probable that certain particles of a nitro-saline nature, and those very subtle, nimble, and of very great fermentative power, are separated from the air by the aid of the lungs and introduced into the mass of the blood. And so necessary for life of every kind is that aereal salt (constituent) that not even plants can grow in earth the access of air to which is shut off. But if that same earth be exposed to air and so forthwith impregnated with that fecundating salt, it at once becomes fit again56 for growing.”17 Mayow fully appreciated the importance of his nitro-aereal particles in the processes of life; he had indeed a fairly accurate conception of a sound theory of animal heat; he saw that they were equally necessary for burning, or combustion, and for respiration, and so was enabled to draw a parallelism between the two processes; he pointed out that they were essential for the ordinary activity of the muscles of the body, that as muscle work was increased more particles from the air were required; indeed, he clearly foresaw the need which the body had for these igneo-aereal particles in all the chemical processes of life. And thus was foreshadowed a conception of oxidation, a hundred years before Priestley evolved his phlogiston theories and Lavoisier discovered oxygen.
The discoveries of Lavoisier, published in 1789, led to a clear understanding of combustion as a process of oxidation, and paved the way for a fuller knowledge of the part played by the oxygen of the air in the chemical reactions going on in the animal body. Lavoisier showed that the oxygen drawn into the lungs with the air breathed was used in the body for the oxidation of certain substances, carbon being transformed thereby into carbon dioxide, and hydrogen into water. Further, he noted that these oxidations were carried forward on a large scale, and he emphasized the importance of oxygen as being the true cause of the varied decompositions taking place in the living body. The larger the amount of oxygen inspired, the more extensive the oxidation, and consequently the rate of respiration as modifying the intake of oxygen served in his opinion as a regulator to control the extent of the oxidative processes. He pointed out that a definite relationship existed between the amount of work done by the body and the oxygen consumed; greater muscular activity, lower temperature of the surrounding air, the activities attending57 the digestive functions, all seemed to be associated with a greater utilization of oxygen. Oxidation was the pivot around which all the chemical reactions of the body seemed to centre. Lavoisier, however, was not a physiologist, and he was, quite naturally perhaps, led into some errors. For example, he considered that the process of combustion or oxidation took place in the lungs, certain fluids rich in carbon and hydrogen formed in the different organs of the body being brought there for exposure to the inspired oxygen. Further, his views implied a simple and complete combustion, in which complex substances rich in carbon were directly and completely oxidized to carbon dioxide and water, in much the same manner as combustion occurs outside the body. Again, he assumed that the amount of oxygen taken into the lungs determined the extent of oxidation, just as the use of the bellows, by increasing the draft of air, causes the fire to burn more brightly.
To Liebig (1842) the next great advance was due. This phenomenally clear-minded man, while recognizing at their full value the fundamental theories advanced by Lavoisier, saw and fully appreciated their incompleteness, and he likewise understood their failure to explain many of the phenomena of life more familiar to the physiological mind than to that of a simple chemist like Lavoisier. Liebig had made a special study of the chemical composition of foodstuffs, and likewise of the tissues and organs of the body. He had, moreover, given great attention to the decomposition products formed in the body, especially the nitrogenous substances excreted through the kidneys, as well as the carbon dioxide and water passed out through the lungs and skin. It was not strange, therefore, that he should take exception to Lavoisier’s view that oxidation in the body consisted in the combustion of a fluid, rich in carbon and hydrogen, which was brought to the lungs. On the contrary, Liebig contended58 that it was the organic compounds, proteids, fats, and carbohydrates, that underwent oxidation, and not necessarily in the lungs, but all over the body, wherever organs and tissues were active. Especially noteworthy was the view advanced by Liebig, and upheld for many years, that of these three classes of compounds the proteids alone served for the construction of organized tissues, like muscle, and that in the activity of this tissue, as in muscle contraction or muscle work, the energy for the work was derived solely from the breaking down or oxidation of this organized proteid. On this ground he termed the proteid foodstuffs “plastic,” or tissue-building foods. Liebig further pointed out that the substances of the body have the power of combining with and holding on to the inspired oxygen, and that fats and carbohydrates, i. e., the non-nitrogenous compounds, easily undergo oxidation or combustion, and thereby furnish the heat of the body. For this reason he termed the corresponding foodstuffs “respiratory” foods. Proteids, on the other hand, according to Liebig’s view, are capable of combustion only in slight degree. The cause of the decomposition of proteid substances in the body was to be traced solely to muscle work, i. e., the energy of muscle contraction, or muscle work, was derived from the breaking down of the proteids of the muscle tissue, and work was the stimulus which brought about proteid decomposition. Non-nitrogenous substances played no part in these reactions; muscle work was without influence on these compounds, oxygen being the sole stimulus which led to their combustion, and heat was the sole product of the combustion.
If Liebig’s theory is correct, that the proteids of the body are decomposed only as the result or the accompaniment of muscle work, and the proteids of the food are used up only as they take the place of the organized proteid so metabolized, it follows that with a like degree of muscular activity a given59 body will always decompose the same amount of proteid. If excess of proteid food is taken, the surplus will be stored in the tissues, or, in other words, the excretion of nitrogen will not be influenced by the amount of proteid consumed in the food. This was the line of argument made use of by various physiologists18 who were disposed to criticise Liebig’s view, and quite naturally the question was soon made the subject of many experiments. It will suffice here merely to say that many concordant results were obtained, showing that an abundance of proteid food leads to an increase in the excretion of nitrogen, muscle activity remaining at a constant level. Hence, as Voit states, some other ground than muscle work must be sought as the true cause of proteid katabolism. Consequently, we find this hypothesis of Liebig replaced by the theory of “luxus consumption,” in which it is maintained that while whatever proteid is used up by the work of the muscle must be made good from the proteid of the food, any excess of proteid absorbed from the intestinal canal is to be considered as “luxus,” and like the non-nitrogenous foods may be burned up in the blood, by the oxygen therein, without being previously organized. Hence, we see suggested two causes for the decomposition of proteid in the body, viz., the work of the muscle and the oxygen of the blood. Further, as stated by C Voit,19 the nitrogen excretion of the hungry or fasting animal affords, according to these views, a measure of the extent to which tissue proteid must be broken down in the maintenance of life, and of the amount of proteid food necessary to be consumed in order to make good the loss; viz., the minimum proteid requirement. Again, since any excess of proteid food beyond this minimal requirement, according to the theory, is destined to be burned60 up in the blood, or elsewhere, to furnish heat the same as non-nitrogenous foods, it follows that the excess of proteid food can be replaced by non-nitrogenous aliment.
Oxidation, however, is the keynote in any explanation of the processes of metabolism, whether nitrogenous or non-nitrogenous matter is involved. Both alike undergo oxidation, but it is not simple oxidation or combustion that we have to deal with. In the time of Lavoisier, as already stated, it was thought that oxygen alone was the cause of the decomposition going on in the body, but simply increasing the intake of air or oxygen, as in quickened breathing or deeper inspiration, does not increase correspondingly the rate of oxidation. In other words, it is not a direct combination of oxygen with the carbon and hydrogen of the foodstuffs, or tissue elements, that takes place in the body, but rather a gradual, progressive decomposition of complex organic compounds into simpler products; made possible, however, by the agency of the oxygen carried from the lungs by the circulating blood. It was demonstrated years ago that animals breathing pure oxygen do not consume any more of the gas than when breathing ordinary air, and likewise no more carbon dioxide is produced in the one case than in the other. Fifty years ago, Liebig and other physiologists showed that frogs’ muscle placed in an atmosphere free of oxygen could be made to contract or do work for some considerable time, and with liberation of heat. This fact implies a breaking down of muscle substance into simpler bodies, but there is here no free oxygen to act as the inciting cause; indeed, what actually occurs is a cleavage or splitting up of substances in the muscle tissue, but at the expense of oxygen in some form of combination in the muscle. This oxygen must have been taken from the blood at some previous time and stored in the tissue for future use. Again, as C Voit has expressed it, if oxygen were really the immediate61 cause of the decompositions taking place in the organism, we should expect combustion to occur in harmony with the well-known relationship of the three classes of organic foodstuffs to oxygen. In other words, fats would undergo combustion most readily, carbohydrates next, and lastly the nitrogenous or albuminous compounds. In reality, however, proteid matter is decomposed in largest quantity; a generous addition of proteid food is always accompanied by an increased consumption of oxygen. Yet oxygen is not the inciting cause of the proteid decomposition, as is seen from the fact that in muscle work, where the intake of oxygen is greatly increased, there is no noticeable change in the amount of proteid material broken down. Plainly, in the body we have to deal not with a direct oxidation of the complex compounds of the tissues or of the food, but rather with a gradual cleavage of these higher compounds into simpler substances, these latter undergoing progressively a still further breaking down with intake of oxygen. To repeat, oxygen is not the cause of the decompositions within the body, but the extent of the breaking down of the tissue or food material is the determining factor in the amount of oxygen taken on and used up. The products of decomposition contain more oxygen than the original substances undergoing the breaking down process, which means that oxygen is taken from the blood and used in the physiological combustion that is going on. It is not, however, strictly a combustion process; it is more complicated and more gradual than ordinary combustion, involving first of all a series of what may be termed oxidative cleavages, in which large molecules are gradually, step by step, broken down into simpler molecules, and these latter then oxidized to still simpler forms. Hence, we find the oxidative changes preceded by a variety of alterations in which oxygen may take no part whatever; such as hydrolytic cleavage, where the elements of water are taken on as a necessary step in the62 cleavage process; dissociation of a simple sort, as when a large molecule breaks up directly into smaller molecules, etc.
These statements by no means detract from the importance of oxygen in the katabolic processes of the body, but it is physiological oxidation that we have to do with, and not simple combustion. Oxygen is not the direct cause of the transformations taking place in the body. As one looks over the history of progress in our knowledge of nutrition from the time of Lavoisier to the present, it is easy to note the gradual change of view regarding oxidation in the living organism. Step by step, it has been demonstrated that there are many factors involved in this breaking down of complex substances; that while oxygen is an ever present requirement, there are other equally important factors to be taken into account. The contrast between the older views and those now current is clearly shown by the difference in attitude regarding the place in the body where oxidation occurs. Thus, in the earlier days, when the view was gradually gaining ground that nutritional changes were mainly the result of oxidation, and that the oxygen drawn into the lungs in inspiration was a primary factor, then, as we have seen, the lungs were considered as the laboratory where the transformation takes place. This view, however, was soon exploded, and next we find the blood, the lymph, and other fluids, but especially the blood, looked on as the locality where oxidation occurs. This was indeed quite a natural view to hold, since the blood is the carrier of oxygen, but we now know, in harmony with the fact that the breaking down of complex food material is a complicated process, involving various kinds of chemical change, that these katabolic processes are not located in any one place, but occur all over the body wherever there are active tissues. As has been previously stated, the human body is a “nation” of cells, all of which are more or less active, and it is in these miniature laboratories mainly that63 oxidation and all the other nutritional changes coincident to life take place. Muscle tissue and nerve tissue, the large secreting glands, such as the liver, stomach, and pancreas, all are the seat of oxidative and other changes which we class under the broad term of nutritional. To these cells, therefore, we must look for an explanation of the causes of oxidation, and the other transformations of a kindred nature that take place in the body.
In our brief survey of digestion, and of the methods there followed for the proper utilization of the organic foodstuffs, it was seen that the unorganized ferments or enzymes are the active agents in accomplishing the breaking down of proteids, and the less profound alteration of fats and carbohydrates. Is it not possible that the tissues of the body are likewise supplied with enzymes of various types, and that upon these powerful agents rests the responsibility for the different kinds of decomposition, oxidation and other changes, that take place in the body? Some years ago much interest was aroused by the observation that certain glands in the body, if simply warmed at body temperature with water, in the presence of some germicidal agent sufficient to prevent putrefactive changes, underwent what is now termed autodigestion, i. e., a process of self-digestion, with formation of various products, notably such as would naturally result from the breaking down of proteid material by ordinary proteolytic enzymes. This would seem to imply the presence in the glands of a proteid-splitting enzyme, the products formed being proteoses, peptones, amino-acids, etc., just such products as result from the action of trypsin. To-day, we know that practically all tissues and organs can, under suitable conditions, undergo autolysis, and in many instances the enzymes themselves can be separated from the tissues by appropriate treatment. Liver, muscle, lymph glands, spleen, kidneys, lungs, thymus, etc., all contain what are very appropriately64 called intracellular enzymes. These enzymes are of various kinds. Especially conspicuous are the hydrolytic, proteid-splitting enzymes, which behave in a manner quite similar to, if not identical with, that of the digestive enzymes of the gastro-intestinal tract, i. e., pepsin, trypsin, and erepsin. Further, there are other hydrolytic cleavages taking place in tissue cells, such as the cleavage of fats, due as we now know to intracellular enzymes of the lipase type, and by which neutral fats are split apart into glycerin and fatty acid. Again, there are in many organs intracellular enzymes which act upon the complex nucleoproteids of the tissue, causing them to break apart into proteid and nucleic acid, the latter being further broken down by other enzymes with liberation of the contained nuclein or purin bases. Many other chemical reactions are brought about by specific enzymes of various kinds, present in the cells of particular glandular organs. Thus, intracellular enzymes have been found, as in the liver, which are able to transform amino-acids into amides, and still others capable of splitting up amides.
Equally important, and even more suggestive, are the data which have been collected recently regarding oxidative processes in the tissues of the body. Specific ferments, known as oxidases, are found widely distributed in many organs and tissues, and it is difficult to escape the conclusion that as intracellular enzymes they have an important part to play in some, at least, of the transformations characteristic of tissue katabolism.20 As a single example, mention may be made of aldehydase, which accomplishes the oxidation of substances having the structure of aldehydes into corresponding acids. Ferments or enzymes of this class are found in the liver,65 spleen, salivary glands, lungs, brain, kidneys, etc., and they may well be considered as important agents in the chemical transformations going on in the tissues of the body. It would take us too far afield to enter into a detailed consideration of these intracellular enzymes; it must suffice to emphasize the general fact that in all the tissues and organs of the body there are present a large number of enzymes of different types, endowed with different lines of activity, and consequently capable of accomplishing a great variety of results in metabolism. Oxidation may still be a dominant feature in nutrition, oxidative changes may characterize more or less every tissue and organ in the body, but the processes are subtle and are not to be defined in harmony with simple chemical or physical laws. The living cell, with its intracellular enzymes, is the guiding and controlling power by which the processes of katabolism are regulated in harmony with the needs of the body. Complex organic matter is broken down step by step in the various tissues, with gradual liberation of the contained energy; processes of hydrolytic cleavage alternate with processes of oxidation, the molecules acted upon growing smaller with each downward step, until at last the final end-products are reached, viz., carbon dioxide, water, and urea, which the body eliminates through various channels as true physiological waste-products.
It will be advisable for us to consider briefly some of these intermediary products of tissue metabolism, since in any discussion of nutritive changes it is quite essential to have some understanding of the chemical relationship existing between the various products which result from the breaking down of proteid and other materials in tissue katabolism. This is especially true of proteid material, since in the gradual disintegration of this substance in tissue metabolism many intermediary bodies are formed, which undoubtedly exercise some physiological influence prior to their transformation66 into simpler bodies, with ultimate formation of the final product, urea. As has been pointed out so many times, the proteid foods are peculiar in that they alone contain the necessary nitrogen, and in the peculiar form able to meet the physiological requirements of the body. Variations in the proteid intake are of necessity accompanied by variations in the formation of nitrogenous intermediary products, and both quality and quantity of these substances must be given due attention in any study of nutrition. Further, it is only by an understanding of the general or ground structure of proteids that we can hope to attain knowledge of the processes going on in the different tissues and organs in connection with metabolism, while a true appreciation of the chemical peculiarities of the individual proteids will help to explain the different nutritional value of vegetable as contrasted with animal proteids.
Our understanding of the chemical structure of any organic substance is based primarily upon a study of the decomposition products which result from its breaking down, under the influence of various chemical agencies. Simple proteid substances when acted upon by pancreatic juice reinforced by the enzyme erepsin, or when boiled with dilute acids, undergo hydrolytic cleavage with ultimate formation of a large number of relatively simple bodies, mostly amino-acids, the chemical structure of which throws some light upon the nature of the proteid. Thus, in the pancreatic digestion of proteid in the intestine we may adopt the following scheme as showing in a general way the progressive transformation that occurs, understanding at the same time that like transformations may be accomplished by corresponding intracellular enzymes in the tissues and organs of the body; and further, that by the long-continued action of hydrolytic agents there is a complete breaking down into amino-acids and other simple products.
67
Native proteid, Protoproteose, Deuteroproteose, Heteroproteose, Primary proteoses, Secondary proteoses, Peptone, Amino-acids
Among these end-products, or amino-acids, are leucin, tyrosin, aspartic acid, glutaminic acid, glycocoll, arginin, lysin, histidin, and likewise the peculiar aromatic body tryptophan. The chemical make-up of these substances may be indicated by the following structural formulæ, which, if even only partially understood, will suggest to the non-chemical mind some idea of close chemical relationship:
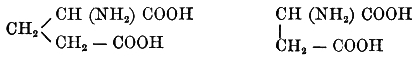
Glutaminic acid Aspartic acid
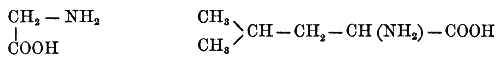
Glycocoll Leucin

Tyrosin
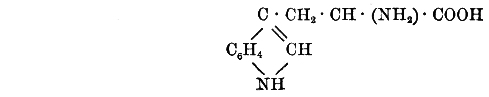
Tryptophan
68
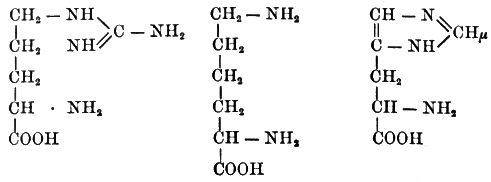
Arginin Lysin Histidin
In these various decomposition products there is apparent certain definite lines of resemblance, on which is based one or more suggestions regarding possible ways in which these chemical groups are linked, or bound together, in the proteid molecule. Thus, there is apparently present a complex or nucleus which may be indicated as

The proteid molecule is presumably built up of amino-acids variously joined together, this synthesis being accomplished, doubtless, by the condensation of different types of amino-acids, in which the first of the above groups represents the more common method of union. We may indeed conjecture that such methods of condensation take place in the human body, in the epithelial cells of the intestine, and in the tissues in general; and that by such methods, construction of proteid is accomplished out of the various fragments split off by digestion, etc. In a tentative way, the principle may be illustrated by the fusion of leucin and glutaminic acid,—following Hofmeister’s suggestion,—in which a still larger complex is formed:
69
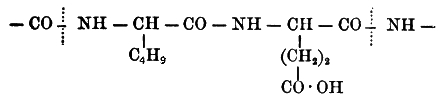
Leucin Glutaminic acid
In this way, step by step, the proteid molecule is built up, and naturally in katabolism the proteid breaks down along certain definite lines of cleavage, with formation of katabolic products containing those groups, or chemical nuclei, which characterize the different proteid molecules. For it is to be clearly understood that there are many different forms of proteid, perhaps superficially alike, but possessed of physiological individuality. This is well illustrated by the two primary proteoses formed in digestion. As will be recalled, there are at first two proteoses produced, protoproteose and heteroproteose. These are, superficially at least, not radically unlike; they possess essentially the same percentage composition, but when broken down by vigorous chemical methods they show a totally different make-up. In other words, at the very beginning of digestion there is a splitting up of the proteid into two parts, which have quite a different chemical structure, as is clearly indicated by the difference in the character and amount of the decomposition products yielded by hydrolytic cleavage. Thus, heteroalbumose as derived from blood-fibrin contains 39 per cent of its total nitrogen in basic form, i. e., in a form which goes over into the basic bodies, arginin, lysin, and histidin, etc. On the other hand, protoalbumose from the same source yields hardly 25 per cent of basic nitrogen. Further, heteroalbumose yields only a very small amount of tyrosin, while protoalbumose gives on decomposition a large amount of this substance. Again, heteroalbumose furnishes a large yield of leucin and glycocoll, while protoalbumose gives no glycocoll and70 only a little leucin. Obviously, these two proteoses have an inner structure quite divergent one from the other, and owing to this fact they must play a quite different rôle in metabolism.
Even greater differences in inner chemical structure are found among native proteids. By way of illustration, we may take egg-albumin, the casein of cow’s milk, gliadin of wheat, and the edestin of hemp seed. These are all typical proteids; they are all useful as food, but they are radically different in their inner chemical structure, as is clearly indicated by the following data,21 which show the percentage yield of the different amino-acids and ammonia:
Leucin. |
Tyrosin. |
Glutam- inic Acid. |
Arginin. |
Lysin. |
Histidin. |
Ammonia. |
|
|---|---|---|---|---|---|---|---|
| Egg-albumin | 6.1 |
1.1 |
9.0 |
. . . |
. . . |
. . . |
1.6 |
| Casein | 10.5 |
4.5 |
10.7 |
4.8 |
5.8 |
2.6 |
1.9 |
| Gliadin | 5.7 |
1.2 |
37.3 |
3.2 |
0 |
0.6 |
5.1 |
| Edestin | 19.9 |
2.7 |
14.0 |
14.2 |
1.6 |
2.2 |
2.3 |
These are not mere technical differences, but they represent divergences of structure which cannot help counting as material factors in nutritional processes. Especially noticeable is the large yield of glutaminic acid from wheat proteid, as contrasted with the proteid (casein) of animal origin. As a rule, glutaminic acid forms a larger proportion of the decomposition products of vegetable than of animal proteids. Similarly, arginin is present in much larger proportion in most vegetable proteids than in most animal proteids. While many other data more or less trustworthy might be added, these figures will suffice to emphasize the main point under discussion, viz., that individual proteids show marked variation71 in the amount of the several amino-acids which serve as corner-stones or nuclei in the building up of the molecule, and consequently they must yield correspondingly different katabolic products when serving the body as food.
Turning now to another phase of tissue metabolism, we may consider briefly the nucleoproteids and their characteristic decomposition products; bodies which are widely distributed as cleavage products formed in the disintegration of most cell protoplasm, and having special interest in nutrition because of their chemical relationship to that well-known substance, uric acid. Nucleoproteids of some type are found in all cells; consequently they are present in all tissues, in all glandular organs, and their widespread distribution constitutes evidence of their great physiological importance. Nucleoproteids are compound substances made up of some form of proteid and nucleic acid. By simple hydrolysis with dilute mineral acids they are broken down into proteid, phosphoric acid, and one or more bodies known as nuclein bases. Of these latter substances, there are four well-defined bodies, viz., adenin, hypoxanthin, guanin, and xanthin, which from their peculiar chemical constitution are known as “purin bases.” In the body, there is present in many cells a peculiar intracellular enzyme termed nuclease, which has the power of liberating these purin bases from their combination as a component part of tissue nucleoproteids, or of the contained nucleic acid. In autolysis or self-digestion of many glands, such as the spleen, thymus, etc., this chemical reaction is easily induced by action of the contained nuclease. Further, the liberated purin bases then undergo change because of the presence of certain deamidizing enzymes, and as a result guanin is transformed into xanthin, and adenin is converted into hypoxanthin. These ferments are true intracellular enzymes, and are termed respectively guanase and adenase. The real essence of the reaction they accomplish is clearly indicated72 by the following formulæ, which likewise show the chemical nature and relationship of the four substances:

Guanin Xanthin

Adenin Hypoxanthin
These two enzymes are typical hydrolyzing enzymes, but it is to be noted that there is not only a taking on of water with a retention of the oxygen, but there is also a giving off of ammonia, by which the transformation is made possible. Adenin is known as an amino-purin and guanin as an amino-oxypurin, while hypoxanthin is an oxypurin and xanthin a dioxypurin. In other words, the two intracellular enzymes are able to transform the two amino-purins into the corresponding oxypurins; i. e., the enzymes are deamidizing ferments, liberating the NH2 group of the adenin and guanin and thus forming two new compounds. These reactions, though more or less technical, are emphasized in this way not merely because they illustrate the action of intracellular enzymes in intermediary metabolism, thus affording a striking example of the gradual changes that take place in ordinary katabolic processes, but especially because they throw light upon the production of another substance common in body metabolism, viz., uric acid. It has long been known that nucleoproteids, nucleins, and other compounds containing73 these purin radicles, when taken as food, cause at once an increased output of uric acid, and it has been clearly recognized that in some way this latter substance, as a product of metabolism, must come from the transformation of nuclein bases. To-day, we understand that in many tissues, as in the liver, spleen, lungs, and muscle, there is present a peculiar oxidizing ferment, an oxidase, by the action of which hypoxanthin can be converted into xanthin, and the latter directly oxidized to uric acid. This conversion into uric acid is purely a process of oxidation, brought about by a typical intracellular oxidase, known specifically as “xanthin oxidase,” the reaction involved being as follows:
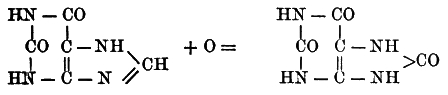
Xanthin Uric acid
From these several reactions, it is clear how various intracellular enzymes working one after the other are able gradually to evolve uric acid from tissue nucleoproteids. Further, it is to be noted that there is another tissue oxidase—contained principally in the kidneys, muscle, and liver—which has the power of oxidizing and thus destroying uric acid, with formation, among other substances, of urea. Remembering that urea has the following chemical constitution
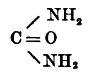
it is easy to see, by comparison of the formulæ, how uric acid might easily yield two molecules of urea through simple oxidation. In this way, excess of uric acid produced in the74 body can be converted into urea, and in this harmless form be excreted from the system.
Finally, reference should be made here to several other products of tissue metabolism, products of the breaking down of proteid matter in the body, since they are liable to prove of interest to us in other connections. Thus creatin, abundant in the muscle and other places; the related substance creatinin, present in the urine; methyl guanidin, a decomposition product of creatin; and urea, all call for a word of description. The chemical relationship of these bodies is clearly indicated by the following formulæ:
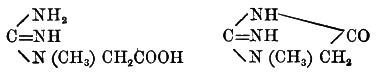
Creatin Creatinin
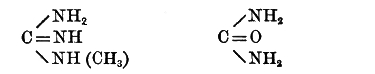
Methyl guanidin Urea
Creatinin is chemically the anhydride of creatin, i. e., it can be formed from creatin by the simple extraction of one molecule of water, H2O. Creatin, by hydrolytic cleavage, will break down into one molecule of urea and one molecule of sarcosin or methyl glycocoll, as shown in the following equation:

Creatin Sarcosin Urea
Methyl guanidin is a decomposition product of creatin, while guanidin, as can be seen from the formula, is like urea, excepting75 that the group NH replaces the oxygen of urea. These simple statements will suffice for our present purpose, viz., to indicate the more or less close chemical relationships existing between many of these nitrogenous decomposition products resulting from proteid katabolism; also to suggest how by slight chemical alteration one decomposition product may be resolved into another related substance in the processes of katabolism. Our conception of the processes involved in proteid katabolism is that of a series of progressive chemical decompositions, in which intracellular enzymes play the all-important part. The intermediary products formed are definite bodies because of the specific nature of the active enzymes, and, secondly, because of the chemical nature of the substances acted upon. In other words, oxidation in the animal body takes the shape of a series of well-defined chemical reactions, in which chemical constitution and specific enzyme action are the predetermining cause. In the absence of the particular chemical groups, the oxidase is unable to bring about oxidation, or, given the proper compound or mother substance in the absence of the specific oxidase, there is no oxidation. Hence, oxidation in the animal body is not the result of simple combustion, but, on the contrary, it consists of a series of orderly chemical processes, each one of which is presided over by an intracellular enzyme, specific in its nature, in that it is capable of acting only upon substances having a certain definite constitution, and leading invariably to a certain definite result. The processes which years ago were considered as due to the peculiar vital properties of the tissue cells, and which were supposed to be entirely dependent upon their morphological and functional integrity, are now seen to be due primarily to a great variety of enzymes, manufactured indeed by the living cells, but capable of manifesting their activity even when free from the influence of the living protoplasm. The varied processes of76 tissue katabolism are the result of orderly and progressive chemical changes, in which cleavage, hydrolysis, reduction, oxidation, deamidization, etc., alternate with each other under the influence of specific enzymes, where chemical constitution and the structural make-up of the various molecules are determining factors in the changes produced.
77
THE BALANCE OF NUTRITION
Topics: Body equilibrium. Nitrogen equilibrium. Carbon equilibrium. Loss of nitrogen during fasting. Influence of previous diet on loss of nitrogen in fasting. Output of carbon during fasting. Influence of pure proteid diet on output of nitrogen. Influence of fat on proteid metabolism. Effect of carbohydrate on nitrogen metabolism. Storing up of proteid by the body. Transformation of energy in the body. Respiration calorimeter. Basal energy exchange of the body. Circumstances influencing energy exchange. Effect of food on heat production. Respiratory quotient and its significance. Influence of muscle work on energy exchange. Elimination of carbon dioxide during work and with different diets. Effect of excessive muscular work on energy exchange. Oxygen consumption under different conditions. Output of matter and energy subject to great variation. Body equilibrium and approximate nitrogen balance to be expected in health.
Man, strictly speaking, is always in a condition of unequilibrium. If placed upon a large and sensitive pair of scales with the opposite side exactly counterpoised, he will be found to lose weight constantly until water or food are taken, when the losses of an hour or two may be made good, or perchance more than balanced. The human body is a maelstrom of chemical changes; chemical decompositions are taking place continuously at the expense of the proteids, fats, and carbohydrates of the tissues and of the food, the stored-up energy of these organic compounds being thereby transformed into the active or “kinetic” forms of78 heat and motion; while carbon dioxide, water, urea, and some few other nitrogenous substances are being continually formed as the normal waste products of these tissue changes, and constantly or intermittently excreted. In other words, the body is in a perpetual condition of chemical oscillation, constantly consuming its own substance, rejecting the waste products which result, and giving off energy in the several forms characteristic of living beings. The condition of the body plainly depends upon the relation which it is able to maintain between the income and the expenditure of matter and energy. If the income equals the output, the body is kept in a condition approaching equilibrium; if the intake exceeds the outgo, the body adds to its capital of matter and energy; while if the expenditure is greater than the income, the accumulated capital is drawn upon; and this, if continued indefinitely, results in a drain upon the bank which must eventually end in disaster. It is comparatively easy, however, for man to maintain his body in a condition of equilibrium from day to day; i. e., the losses of the morning can be made good at luncheon, or the expenditures of an entire day counterbalanced by a corresponding addition to capital the following day, in which case the body may be said to be in balance. It is necessary, however, to discriminate between body equilibrium, meaning thereby the maintenance from day to day of a constant body-weight, and nitrogen equilibrium, or carbon equilibrium. In the latter cases, what is meant is that the intake of nitrogen, or of carbon, exactly equals the output of these two elements. It is quite possible, however, to have a condition of nitrogen equilibrium without the body being in a state of balance, as when the outgo of carbon exceeds the intake of carbon, or when there is an increased output of water.
As a rule, it may be stated that when a man puts out less carbon and less nitrogen than he takes in he must be gaining79 in weight; the only exception being the possible case of an increased excretion of water, which might more than counterbalance the gain. On the other hand, if he gives off more carbon and more nitrogen than he takes in, the body must lose in weight. Where the output of carbon is beyond the amount of carbon ingested, the lost carbon represents a drain upon body fat. In a reversal of this condition, i. e., where the carbon taken in is in excess of the outgo, the body is gaining in fat. Theoretically, gain or loss of carbon may mean gain or loss of either carbohydrate or fat, but practically stored-up carbon generally stands for accumulated fat; and, correspondingly, loss of carbon represents a withdrawal from the store of adipose tissue, since glycogen and sugar from a quantitative standpoint figure only slightly in these metabolic processes. When the body excretes more nitrogen than is taken in during a given period, there is only one interpretation possible, viz., that the body is losing proteid or flesh. If, on the other hand, the nitrogen import exceeds the outgo, then the body must be gaining flesh. Here, again, there is the theoretical possibility that gain or loss of nitrogen might represent increase or decrease of proteid in some glandular organ, or even in the blood; but practically it is the relatively bulky muscle tissue, with its high content of proteid matter, that is most subject to change in metabolism. Finally, it is easy to see how, knowing the percentage of nitrogen in proteid and the percentage of carbon in fat, one can calculate from the nitrogen and carbon lost or gained the amounts of proteid or fat added to the capital stock, or withdrawn from the store of nutritive material.
When there is no income, as in fasting, the body loses rapidly, living during the hunger period upon its store of energy-containing material. Many careful observations have been made upon people who have fasted for long periods, some as long as thirty days, the income consisting solely of80 water. The following figures22 show the daily excretion of nitrogen in several notable cases:
Day of Fasting. |
Breithaupt. 59.9 Kilos. |
Cetti. 56.5 Kilos. |
Succi. 62.4 Kilos. |
|---|---|---|---|
grams |
grams |
grams |
|
0 |
13.0 |
13.5 |
16.2 |
1 |
10.0 |
13.6 |
13.8 |
2 |
9.9 |
12.6 |
11.0 |
3 |
13.3 |
13.1 |
13.9 |
4 |
12.8 |
12.4 |
12.8 |
5 |
11.0 |
10.7 |
12.8 |
6 |
9.9 |
10.1 |
10.1 |
7 |
. . . |
10.9 |
9.4 |
8 |
. . . |
8.9 |
8.4 |
9 |
. . . |
10.8 |
7.8 |
10 |
. . . |
9.5 |
6.7 |
In Succi’s case, the fasting was continued for thirty days. The daily average loss of nitrogen from the 11th to the 15th day was 5.8 grams; from the 16th to the 20th day, 5.3 grams; from the 20th to the 25th day, 4.7 grams; and from the 26th to the 30th day, 5.3 grams. A daily loss of 5.3 grams of nitrogen means a breaking down, or using up, of 33 grams of proteid, or a little more than one ounce. On the sixth day of fasting, all three of these subjects showed essentially the same daily loss of nitrogen; viz., 10 grams, which implies a using up of 62.5 grams of proteid material. We must not be led astray by these figures, however, or draw too hasty81 conclusions therefrom regarding the requirements of the body for proteid food. Noting the close agreement in the nitrogen output of the three subjects on the sixth day, combined with the fact that their body-weight was essentially the same, we might infer that 62.5 grams of proteid matter represents the amount of nitrogenous food necessary to maintain nitrogen equilibrium and keep the body in a condition of balance. Such a conclusion, however, would be quite erroneous for several reasons. First, a man fasting, if he was in an ordinary condition of nutrition prior to the fast, has in his tissues a large store of fat. It is considered that in fasting only about 10–12 per cent of the total energy of the body is derived from tissue proteid; the major part comes from the fat stored up. When there is no income to make good the loss, the body must naturally draw upon its own store. A certain amount of proteid must be used up daily, but in addition there are the energy requirements to be considered. These are met mainly by fat and carbohydrate, and so long as fat endures proteid will be drawn upon only, or mainly, to meet the nitrogen requirement; but if the fat gives out, then proteid must be used in larger quantity, as a source of energy. Hence in fasting, the daily loss of nitrogen will be governed largely by the condition of the body as regards fat. Thus, Munk has reported the case of a well-nourished and fat person, suffering from disease of the brain, who gave off daily in the later stages of starvation only one-third the amount of nitrogen voided by Cetti, who had been poorly nourished. Obviously, in fasting, as soon as the adipose tissue of the body has been largely used up, there will be an increase in the amount of tissue proteid consumed, since under such conditions the heat of the body and the energy of muscular work (work of the heart and involuntary muscles) must come from the decomposition of proteid. In harmony with this statement, it is frequently observed that in cases of starvation82 there comes toward the end a sudden and marked increase in the output of nitrogen.
Secondly, the elimination of nitrogen during the earlier days of fasting is governed in large measure by the character and extent of the diet on the days just preceding the fast. This is well illustrated by some experiments conducted by C Voit on a dog. In the first series of experiments, the dog received daily 2500 grams of meat prior to fasting; in the second series, 1500 grams of meat were fed daily before the fast; while in the third series, a mixed diet relatively poor in proteid was given. The following figures23 show the amounts of proteid used up by the dog (calculated from the nitrogen excreted) each day of the fasting period, under the different conditions:
First Series. |
Second Series. |
Third Series. |
|
|---|---|---|---|
grams |
grams |
grams |
|
| First fasting day | 175 |
77 |
40 |
| Second " " | 72 |
54 |
33 |
| Third " " | 56 |
46 |
30 |
| Fourth " " | 50 |
53 |
36 |
| Fifth " " | 36 |
43 |
35 |
| Sixth " " | 39 |
37 |
37 |
We see very clearly in these experiments the effects of the large quantities of proteid fed on the destruction of proteid in the early days of fasting. When the body is rich in proteid from food previously taken, the metabolism of nitrogenous matter is very large at first, as in the first series of experiments. Indeed, in this series, even on the fifth day of83 fasting, the amount of proteid metabolized was larger than on the second day of the third series. We have here a forcible illustration of the physiological axiom that excess of proteid matter in the tissues, or in the blood, stimulates proteid metabolism; and it affords convincing proof of the contention that in the first days of fasting the output of nitrogen, or the amount of proteid used up, will depend in large measure upon the proteid condition of the body at the time of the fast. Equally noticeable is the fact that there comes a time—the sixth day in the above experiment—when the nitrogen output reaches a common level, irrespective of the previous proteid condition of the body. Further, it is easy to see that the greater loss of nitrogen, i. e., the large breaking down of proteid during the first few days of fasting, in those cases where proteid food has been freely taken, suggests the existence in the tissues of two forms of proteid. We may term them, following the nomenclature of Voit, as circulating and morphotic, or tissue, proteid; or, we may designate them as labile and stable forms of proteid. In other words, following the usually accepted view, this circulating or labile proteid represents reserve or surplus material which is easily decomposed and hence rapidly gotten rid of, while the stable proteid is more slowly oxidized, and its metabolism may be taken as representing more nearly the real necessities of the body. However this may be, it is plainly manifest that the nitrogen output, meaning the metabolism of proteid matter, during hunger or fasting is modified by a variety of circumstances, notably the previous nutritive condition of the body as regards both fat and proteid. It is hardly necessary to add that the amount of muscular work performed is another factor of importance in this connection. Fat in the body represents inert material stored up mainly for nutritive purposes; hence, in hunger it is used largely, and serves to protect more important tissues. Thus, experiments have shown84 that in long periods of fasting, adipose tissue may be consumed to the extent of 97 per cent of the total amount present, while the heart and nervous tissue will not lose over 3 per cent of their tissue substance. The influence of tissue fat upon the consumption of proteid during hunger can thus be fully appreciated.
The output of carbon during fasting may be illustrated by the following experiment24 made upon a young man, the nitrogen data being included for comparison, and likewise the intake of food, in terms of nitrogen and carbon, preceding the fast and for two days following the fast. The fasting was of five days’ duration.
Day. |
Body-weight. |
Intake. |
Output. |
||
|---|---|---|---|---|---|
Carbon. |
Nitrogen. |
Carbon.25 |
Nitrogen. |
||
kilos |
grams |
grams |
grams |
grams |
|
2 |
67.4 |
438.7 |
30.96 |
303.4 |
25.81 |
3 |
66.9 |
0 |
0 |
197.6 |
12.17 |
4 |
65.7 |
0 |
0 |
188.8 |
12.85 |
5 |
64.8 |
0 |
0 |
183.2 |
13.61 |
6 |
63.9 |
0 |
0 |
180.8 |
13.69 |
7 |
63.1 |
0 |
0 |
176.2 |
11.47 |
8 |
63.9 |
439.9 |
35.65 |
270.5 |
26.83 |
9 |
65.5 |
391.7 |
23.68 |
258.8 |
19.46 |
On the non-fasting days, the intake consisted of an ordinary food mixture of proteids, fats, and carbohydrates, with a small addition of alcohol. The point to be emphasized85 here, however, is that the carbon-content was more than sufficient to meet the needs of the body. Thus, it will be observed that on all three of the days when food was taken, the income of carbon was far in excess of the output. In other words, on the day preceding the beginning of the fast the body stored up 135 grams of carbon, and on the day following the fast the body retained 169 grams of carbon to help make good the loss. Similarly, the amount of proteid food taken in on the day prior to the fast was considerably in excess of the needs of the body, 5.1 grams of nitrogen equivalent to 31.8 grams of proteid being stored for future use. Plainly, the man was not in either carbon or nitrogen balance prior to the fast, but was taking far more food than the needs of the body called for. This fact may be emphasized by noting that the total fuel value of the daily food, plus the fuel value of the alcohol, amounted on an average to about 4200 large calories, while the fuel value of the material metabolized on the feeding days averaged only 2500 calories. Looking at the figures showing the output of carbon, as well as of nitrogen, during the fasting days, it is to be seen that in the early days of fasting, the metabolism of the body tends to remain at a fairly constant level, especially when figured per kilogram of body-weight.
To fully appreciate what takes place in a man of the above body-weight fasting for five days (though living on a large excess of food prior to the fast), the daily losses of carbon and nitrogen may be translated into terms of fat and proteid. If it is assumed that the total carbon, aside from what necessarily belongs to the proteid indicated by the nitrogen figures, comes from the oxidation of fat, it is easy to compute the amounts of fat and proteid metabolized, or destroyed, each day of the fasting period. These are shown in the following table:
86
Day. |
Proteid metabolized. |
Fat metabolized. |
|---|---|---|
grams |
grams |
|
3 |
76.1 |
206.1 |
4 |
80.3 |
191.6 |
5 |
85.1 |
181.2 |
6 |
85.6 |
177.6 |
7 |
71.7 |
181.2 |
Finally, if from these figures we calculate the fuel value of the proteid and fat oxidized per day, it is possible to gain a fairly clear conception of the part played by these two classes of tissue material during fasting, in furnishing the heat of the body and the energy for muscular motion, etc.
Day. |
Fuel Value of the Proteid metabolized. |
Fuel Value of the Fat metabolized. |
Total Fuel Value. |
|---|---|---|---|
calories |
calories |
calories |
|
3 |
303 |
1916 |
2220 |
4 |
320 |
1781 |
2102 |
5 |
339 |
1684 |
2024 |
6 |
341 |
1651 |
1992 |
7 |
286 |
1684 |
1970 |
These somewhat general statements, with the illustrations given, will serve in a brief way to emphasize some of the essential features of metabolism in the fasting individual; where there is no income of energy-containing material, and where the body must draw entirely upon its store of accumulated fat and proteid to keep the machinery in motion, maintain body temperature, and do the tasks of every-day life. When it is remembered that persons have fasted for periods87 of thirty days or longer without succumbing, it is evident that the body of the well-nourished man has a large reserve of nutritive material, which can be drawn upon in cases of emergency. At the same time, the facts presented show us that in the early days of fasting the actual amounts of tissue proteid and body fat consumed are not large. In Cetti’s case, on the sixth day of fasting the metabolized nitrogen amounted to 10 grams, which implies a loss of 62.5 grams of proteid. At this rate of loss, one pound of dry proteid matter in the form of tissue proteid would meet the wants of a man of 130 pounds body-weight for seven and a half days, provided of course there was a reasonable stock of fat to help satisfy the energy requirements. Finally, we may again emphasize the fact that the loss of nitrogen in the fasting man is by no means a measure of the minimal proteid requirement. By feeding fat, or carbohydrate, or both, the output of nitrogen can be materially diminished, although naturally we cannot establish a nitrogen balance by so doing, since the income is free from nitrogen; but we can postpone for a time the approach of nitrogen starvation.
We may next profitably consider the effect of a pure proteid diet—such as lean meat free from fat—on the output of nitrogen. In studying this problem, we at once meet with several important and surprising facts. First, we are led to see that, strange as it may seem, every addition of proteid to the diet results in an increased excretion of nitrogen. In other words, increase of proteid income is followed at once by an increase in the metabolism of proteid, with a corresponding outgo of nitrogen. The hungry or fasting man with his income entirely cut off, and consequently suffering from a heavy drain upon his capital stock, would be expected, when suddenly supplied with fresh capital in the form of meat or other kind of proteid food, to hold on firmly to this all-important foodstuff; but such is not the case. It is impossible,88 for example, to establish nitrogen equilibrium by an income of proteid equal to what the individual during fasting is found to metabolize. As stated by another, “It is one of the cardinal laws of proteid metabolism that the store of nitrogenous substances in the body is not increased by, or not in proportion to, an increase in the nitrogen intake.” The principle is well illustrated in the fasting experiment just described. On the fifth day of fasting, the nitrogen output amounted to 11.4 grams. On the day following, the man took 35.6 grams of nitrogen in the form of proteid, while the excretion of nitrogen for that day rose to 26.8 grams. In other words, although deprived of all proteid income for five days, and during that period drawing entirely upon his proteid capital, the man was wholly unable to avail himself of the proteid so abundantly supplied at the close of the fast and make good the losses of the preceding days; only a small proportion of the proteid income could be retained. If a dog fed on a definite quantity of meat suddenly has his proteid income increased, there is at once an acceleration of proteid metabolism, and a corresponding increase in the output of nitrogen. Addition of still more proteid to his income is followed by an accumulation of a portion of the proteid; but this tends to decrease gradually, while there is a corresponding daily increase in the excretion of nitrogen. In this manner, there finally results a condition of nitrogenous equilibrium or nitrogen balance.
Again, an animal brought into nitrogen equilibrium by excessive proteid feeding, if suddenly given a small amount of meat per day, tends to put out nitrogen from its own tissues. This tissue loss, however, decreases slowly, and eventually the animal is quite likely to re-establish nitrogen equilibrium at a lower level. There is, in other words, a strong tendency for the body to pass into a condition of nitrogen balance under different conditions of proteid89 feeding, even after a long period of nitrogen loss and with an abundance of proteid in the intake. The starving body, as we have seen, cannot make use of all the nitrogen fed, although we can well conceive its great need for all the proteid available. A certain amount of the proteid fed, or its contained nitrogen, is at once passed out of the body and lost, even though the organism be gasping, as it were, for proteid to make good the drain incidental to long fasting. A recent writer26 has suggested that some explanation for these anomalies may be found in the supposition “that a long succession of generations in the past, which have lived from choice or necessity on a diet rich in proteids, have handed down to us, as an inheritance, a constitution in which arrangements exist for the removal of nitrogen from a considerable part of this proteid. The fact that the amount of proteid taken is re-adjusted to suit the actual needs of the body, though it makes these arrangements unnecessary, will not necessarily remove them. The denitrifying enzyme, which has been trained to keep guard over the entrances by which nitrogenous substances are admitted into the body, will continue to levy its toll of nitrogen, even when the amount of proteid presented to it is no more than the tissues which it serves actually require.”
As an illustration of how the body behaves with a low nitrogen intake followed by a sudden increase in the income of proteid, some data from an experiment performed by Sivén27 on himself may be cited:
90
Date. |
Body-weight. |
Nitrogen of the Food. |
Nitrogen excreted. |
Nitrogen Balance. |
|---|---|---|---|---|
kilos |
grams |
grams |
grams |
|
Nov. 6 |
65.4 |
2.69 |
8.31 |
–5.62 |
7 |
65.4 |
2.69 |
5.37 |
–2.68 |
8 |
65.1 |
2.69 |
5.71 |
–3.02 |
9 |
65.3 |
2.69 |
4.88 |
–2.19 |
10 |
65.0 |
2.69 |
4.32 |
–1.63 |
11 |
64.9 |
2.69 |
4.25 |
–1.56 |
12 |
64.9 |
2.69 |
4.47 |
–1.78 |
13 |
64.6 |
2.96 |
4.88 |
–1.92 |
14 |
64.4 |
2.96 |
4.30 |
–1.44 |
15 |
64.3 |
2.96 |
4.75 |
–1.79 |
16 |
64.4 |
2.96 |
4.36 |
–1.40 |
17 |
64.4 |
2.96 |
4.13 |
–1.17 |
18 |
64.4 |
2.96 |
4.35 |
–1.39 |
19 |
64.4 |
2.96 |
4.32 |
–1.36 |
20 |
64.4 |
2.96 |
4.22 |
–1.26 |
21 |
64.0 |
2.96 |
4.06 |
–1.10 |
–31.31 |
||||
22 |
64.1 |
4.02 |
4.22 |
–0.20 |
23 |
64.4 |
4.02 |
4.35 |
–0.33 |
24 |
64.4 |
4.02 |
4.21 |
–0.19 |
25 |
64.4 |
4.02 |
4.40 |
–0.38 |
–1.10 |
||||
26 |
64.2 |
8.24 |
6.56 |
+1.68 |
27 |
64.4 |
13.45 |
8.67 |
+4.78 |
28 |
64.4 |
13.66 |
10.54 |
+3.12 |
29 |
64.0 |
13.45 |
11.10 |
+2.35 |
30 |
64.2 |
13.24 |
12.83 |
+0.41 |
Dec. 1 |
64.2 |
13.24 |
11.70 |
+1.54 |
2 |
63.9 |
12.61 |
12.00 |
+0.61 |
+14.49 |
||||
3 |
64.0 |
22.93 |
16.24 |
+6.69 |
4 |
63.9 |
22.41 |
21.47 |
+0.94 |
5 |
63.9 |
22.41 |
23.10 |
–0.69 |
6 |
63.6 |
23.35 |
23.12 |
+0.23 |
7 |
63.9 |
23.04 |
22.82 |
+0.22 |
8 |
63.8 |
22.62 |
22.86 |
–0.24 |
+6.15 |
91
I have ventured to give these data in some detail, because of their exceeding great interest in several directions aside from the point under discussion. Confining our attention to the nitrogen exchange, it is to be observed that for a period of two weeks Sivén lived on less than 3 grams of nitrogen per day, and without any excessive intake of carbohydrate or fat. During this time, the body naturally was in a condition of minus balance as regards nitrogen, the output being considerably larger than the income. The total amount of nitrogen lost in the period, 31 grams, corresponds to a breaking down of 193 grams of tissue proteid, or over one-third of a pound. On increasing the income of nitrogen to 4 grams per day, the nitrogen loss still continued, though at a much lower rate; indeed, the body is seen to approach very closely to a condition of nitrogen equilibrium. Still further increase of the nitrogen income to 13 grams per day was followed at once by a slight accumulation of proteid, and the body showed a decided plus balance of nitrogen, as on November 27. This, however, is seen to decrease gradually with a corresponding daily increase in the outgo of nitrogen, until on December 2 the body was once more practically in nitrogenous equilibrium. On again increasing the nitrogen income, to 23 grams per day, the same process was repeated, although in this case the body more quickly approached a condition of nitrogen balance.
We see in these data striking confirmation of the statement that the nitrogen outgo tends to keep pace with the income of nitrogen, the body always striving to maintain a condition of nitrogen equilibrium. Consequently, the fasting man having lost largely of his store of proteid can replace the latter only slowly, even though he eats abundantly of proteid food. Thus, Sivén in the week ending December 2, though taking over 13 grams of nitrogen a day, retained in his body only 14.5 grams of nitrogen during the entire seven days; while in the six days following, with a daily intake of 23 grams of nitrogen,92 he gained only about 8 grams additional. The human body does not readily store up proteid, and this is true no matter how greatly the tissues are in need of replenishment.
If the daily income is reinforced by the addition of carbohydrate or fat, there is observed a decided influence on the outgo of nitrogen; the rate or extent of proteid metabolism is at once modified, fat and carbohydrate both having a direct saving effect on proteid. Neither fat nor carbohydrate can prevent the katabolism of proteid, but they can and do decrease it, and thus serve as proteid-sparers. In the fasting body, or where there is only an intake of proteid, the latter material, except for the fat contained in the tissues, must serve the double purpose of meeting the specific nitrogen requirements of the body and furnishing the requisite energy. The energy requirements, however, can be met more advantageously by either of the non-nitrogenous foodstuffs, and just so far as they are oxidized, so far will there be a saving of proteid. Herein lies the philosophy of a mixed diet, with its natural intermingling of proteid, fat, and carbohydrate. For the same reason, the body of a man rich in fat will in fasting lose far less proteid per day than the lean man; or, if fed with a given amount of proteid food, the fat man may attain nitrogen equilibrium, or even store up a little proteid, while on the same diet the lean man will lose proteid. Further, if a man is in nitrogen balance with a given amount of proteid food, the addition of fat or carbohydrate to the diet will permit of a reduction in the amount of proteid necessary to maintain nitrogenous equilibrium. Fat, however, when added to food, does not always protect proteid to the extent possibly suggested by the preceding statements. The following data from oft-quoted experiments by Voit28 on dogs will serve to illustrate:
93
Food. |
Flesh. |
||
|---|---|---|---|
Meat. |
Fat. |
Metabolized. |
On the Body. |
grams |
grams |
grams |
grams |
1500 |
0 |
1512 |
–12 |
1500 |
150 |
1474 |
+26 |
500 |
0 |
556 |
–56 |
500 |
100 |
520 |
–20 |
Food. |
Flesh. |
||
|---|---|---|---|
Meat. |
Fat. |
Metabolized. |
On the Body. |
grams |
grams |
grams |
grams |
1500 |
0 |
1512 |
–12 |
1500 |
150 |
1474 |
+26 |
500 |
0 |
556 |
–56 |
500 |
100 |
520 |
–20 |
It is to be observed that in both of these experiments the fairly large addition of fat results in a saving of proteid, but the sparing effect in the first experiment amounts to only 38 grams of proteid for the 150 grams of fat added. In the second experiment, however, there is a saving of 36 grams of proteid, although only 100 grams of fat were fed. The radical point of difference in the two experiments is the amount of proteid ingested. Proteid food stimulates proteid metabolism; it likewise accelerates the metabolism of non-nitrogenous matter, consequently the sparing or protecting effect of fat on proteid is most conspicuous when the intake of proteid is relatively small. Only under such conditions, does fat protect in large degree the consumption of proteid in the body. In the ordinary, daily, dietary of man, with its great variety of food materials and with its proteid-content not exceeding 125 grams, fat is apt to be a conspicuous element, and under such conditions its sparing effect on proteid metabolism is most marked. Further, it must not be forgotten, as Voit originally pointed out, that the adipose tissue of the body acts like the food-fat, and consequently the proteid-sparing effect of the former may be added to that of the latter.
The addition of carbohydrate to a meat diet produces at94 once a saving in the decomposition of proteid, as shown in the following figures, covering an experiment of two days:
Meat. |
Sugar. |
Proteid metabolized. |
| 500 grams. | 200 grams. | 502 grams. |
| 500 | 0 | 564 |
Without the sugar, there was a minus balance of 64 grams of proteid, but addition of the carbohydrate caused practically a saving of all of this, with establishment of essentially a nitrogen balance. The sparing of proteid by carbohydrate is greater than by fats, a fact of considerable dietetic importance which is well illustrated by the following experiments (on dogs) taken from Voit:
Food. |
Flesh. |
||
|---|---|---|---|
Meat. |
Non-nitrogenous Food. |
Metabolized. |
Balance of the Body. |
grams |
grams |
grams |
grams |
500 |
250 Fat | 558 |
–58 |
500 |
300 Sugar | 466 |
+34 |
500 |
200 Sugar | 505 |
–5 |
800 |
250 Starch | 745 |
+55 |
800 |
200 Fat | 773 |
+27 |
2000 |
200–300 Starch | 1792 |
+208 |
2000 |
250 Fat | 1883 |
+117 |
Food. |
Flesh. |
||
|---|---|---|---|
Meat. |
Non-nitrogenous Food. |
Metabolized. |
Balance of the Body. |
grams |
grams |
grams |
grams |
500 |
250 Fat | 558 |
–58 |
500 |
300 Sugar | 466 |
+34 |
500 |
200 Sugar | 505 |
–5 |
800 |
250 Starch | 745 |
+55 |
800 |
200 Fat | 773 |
+27 |
2000 |
200–300 Starch | 1792 |
+208 |
2000 |
250 Fat | 1883 |
+117 |
In considering the results of this experiment, it must be remembered that the calorific or fuel value of fat as compared with carbohydrate is as 9.3 : 4.1; in other words, fats have a fuel value of more than twice that of carbohydrates. In spite of this fact, it is clearly evident that carbohydrates as a class—for the different sugars and starches act alike in this respect—are far more efficient than fats in saving proteid. Thus, with an income of 500 grams of meat and 25095 grams of fat, the body of the animal lost 58 grams of proteid, while with a like amount of meat and 300 grams of sugar the body not only saved the 58 grams, but in addition stored 34 grams of proteid, showing a plus balance to that extent. The sparing of proteid by carbohydrate amounts on an average, according to Voit, to 9 per cent—in the highest cases to 15 per cent—of the proteid given, while the saving produced by fat averages only 7 per cent. Further, increasing quantities of carbohydrates in the food diminish the rate of proteid metabolism much more regularly and constantly than increasing quantities of fat. We may attribute this difference in action, in a measure at least, to the greater ease in oxidation and utilization of the carbohydrate. In any event, starches and sugars are most valuable adjuncts to the daily diet, because of this marked proteid-saving power, while their fuel value adds just so much to the total energy intake.
A more striking illustration of the action of carbohydrate in sparing proteid is seen in experiments on man, where the nitrogen intake is reduced to a minimum, so as to constitute a condition of specific nitrogen-hunger. In such a case, increasing amounts of carbohydrate added to the intake reduce enormously the using up of tissue proteid. The following experiment with a young man 22 years old and 71.3 kilos body-weight, reported by Landergren,29 affords good evidence of the extent to which this proteid sparing power may manifest itself.
We see here the nitrogen consumption fall to the exceedingly low level of 3.34 grams per day, or 0.047 gram per kilo of body-weight. To appreciate the full significance of this drop in the extent of proteid metabolism, we may recall that Succi, with a body-weight of only 62.4 kilos, on the seventh day of fasting excreted 9.4 grams of nitrogen, corresponding96 to a metabolism of 58.7 grams of tissue proteid. In other words, with an intake of only 5.6 grams of proteid, the addition of 908 grams of carbohydrate, with a total fuel value of 3745 calories, reduced the consumption of tissue proteid to 20.8 grams. The same individual, if fasting, would undoubtedly have used up at least 70 grams of tissue proteid.
Day. |
Intake. |
Output. |
Proteid metabolized. |
||||
|---|---|---|---|---|---|---|---|
Proteid. |
Fat. |
Carbo- hydrate. |
Alcohol. |
Calories. |
Nitrogen of Urine. |
||
grams |
grams |
grams |
grams |
grams |
grams |
||
1 |
35.2 |
6.1 |
507 |
26.6 |
2465.9 |
12.16 |
76.0 |
2 |
28.7 |
4.7 |
787 |
26.6 |
3574.3 |
8.37 |
52.3 |
3 |
28.8 |
4.7 |
841 |
26.6 |
3796.1 |
5.02 |
31.3 |
4 |
28.3 |
4.9 |
839 |
13.3 |
3690.5 |
4.50 |
28.1 |
5 |
5.4 |
. . |
898 |
. . . . |
3703.9 |
4.01 |
25.0 |
6 |
6.0 |
. . |
931 |
. . . . |
3841.7 |
3.36 |
21.0 |
7 |
5.6 |
. . |
908 |
. . . . |
3745.8 |
3.34 |
20.8 |
It is evident from what has been said that both of these non-nitrogenous foods, fat and carbohydrate, play a very important part in nutrition, because of their ability to maintain in a measure the integrity of tissue proteid. When we recall that a diet of pure proteid, such as meat or eggs, must be excessive in quantity in order to meet the energy requirements of the body, and that the stimulating action of proteid food serves to whip up body metabolism, we can appreciate at full measure the great physiological economy which results from the addition of carbohydrate and fat to the daily diet. The establishment of nitrogenous equilibrium is made possible at a much lower level by the judicious addition of these two non-nitrogenous foodstuffs. The same principle may be illustrated in another way, viz., by noting the effect on tissue97 proteid of withdrawal of a portion of the fat or carbohydrate of the intake, in the case of a person nearly or quite in nitrogen balance. The following experiment30 affords a good example of what will occur under such treatment:
Income. |
Output of Nitrogen. |
Balance of Nitrogen in Body. |
||||
|---|---|---|---|---|---|---|
Nitrogen. |
Fat. |
Carbo- hydrate. |
Calories. |
|||
grams |
grams |
grams |
grams |
|||
Av. of 3 days |
15.782 |
40.47 |
289.6 |
1955 |
14.927 |
+0.862 |
Nov. 30 |
15.782 |
40.34 |
177.3 |
1493 |
14.959 |
+0.830 |
Dec. 1 |
15.782 |
40.34 |
177.3 |
1493 |
17.546 |
–1.757 |
2 |
15.782 |
40.34 |
177.3 |
1493 |
18.452 |
–2.663 |
Average of the last two days . . . . . . . . . -2.210 |
||||||
Starting with the body in a condition of plus nitrogen balance, i. e., with a mixed diet more than sufficient to maintain the tissue proteid intact, the reduction of the fuel value of the food from 1955 to 1493 calories by cutting off 112 grams of carbohydrate per day was followed by a gradual, but marked, increase in the output of nitrogen; indicating thereby the extent to which the body proteid was then drawn upon to make good the loss of energy-containing income. The body showed at the close of the experiment a minus nitrogen balance averaging 2.2 grams per day, or a loss of 13.8 grams of tissue proteid, which would obviously have continued, under the above conditions, until the body was exhausted. In other words, the 112 grams of carbohydrate, if added to the diet on December 3 and the following days, would have quickly saved the daily loss of 2.4 grams of nitrogen, and thus changed the drain of tissue proteid to an98 actual gain, with consequent establishment of a growing plus balance.
It is obvious from what has been stated, that in man the body can accomplish a storing of proteid only when the intake is reinforced by substantial additions of fat or carbohydrate. It is plainly a matter of great physiological importance that the body should be able to increase at times its reserve of proteid. This, however, cannot apparently be accomplished on a large scale under ordinary conditions. Any storing up of nutritive material in excess, whether it be proteid or fat, necessarily involves overfeeding, i. e., the taking of an amount of food beyond the capacity of the body to metabolize at the time. Fat, as we know, may be stored in large quantities, and it is in cases of overfeeding with non-nitrogenous foods that we find accumulation of fat most marked. Overfeeding with proteid, however, does not lead to corresponding results, owing primarily to the peculiar physiological properties of proteid; its general stimulating effect on metabolism, the tendency of the body to establish nitrogenous equilibrium at different levels, and the fact emphasized by von Noorden that flesh deposition is primarily a function of the specific energy of developing cells. In other words, the protoplasmic cells of the body are more important factors in the storing or holding on to proteid than an excess of proteid-containing food.
It is generally considered as a settled fact, that in man it is impossible to accomplish any large permanent storing or deposition of flesh by overfeeding. Similarly, it is understood that the muscular strength of man cannot be greatly increased by an excessive intake of food. The only conditions under which there is ordinarily any marked and permanent flesh deposition are such as are connected with the regenerative energy of living cells. Thus, as von Noorden has stated, an accumulation or storing of tissue99 proteid is seen especially in the growing body, where new cells are being rapidly constructed; also in the adult where growth may have ceased, but where increased muscular work has resulted in an hypertrophy or enlargement of the muscular tissue; and lastly in those cases where, owing to previous insufficient food or to the wasting away of the body incidental to disease, the proteid content of the tissues has been more or less diminished, and consequently an abundance of proteid food is called for and duly utilized to make good the loss. In some oft-quoted experiments by Krug, conducted on himself, it was observed that with an abundant food intake, sufficient to furnish 2590 calories per day (44 calories per kilo of body-weight), a condition approaching nitrogenous equilibrium was easily maintained. On then increasing the fuel value of the food to 4300 calories (71 calories per kilo of body-weight) by addition of fat and carbohydrate, there was during a period of fifteen days a sparing of 49.5 grams of nitrogen or 309 grams of proteid, which would correspond to about 1450 grams, or three pounds, of fresh muscle. It is to be noted, however, that of this excess of calories added to the intake only 5 per cent was made use of for flesh deposit, the remaining 95 per cent going to make fat.
Again, we may call attention to the well-known fact that in feeding animals for food, while fat may be laid on in large amounts, flesh cannot be so increased by overfeeding. In this matter, however, race and individuality count for considerable. Thus, there is on record a more recent series of experiments conducted by Dapper31 on himself which shows some remarkable results. Starting with a daily diet not excessive in amount, he was able by an addition of only 80 grams of starch to accomplish a laying up of 3.32 grams of nitrogen100 per day for a period of twelve days, or a total gain of 39.8 grams of nitrogen, equal to 248 grams of proteid. It may be said that the gain of proteid or flesh here for the twelve days was no greater than in the preceding case (fifteen days), but the difference lies in the fact that Krug accomplished his gain by increasing the daily intake from 2590 to 4300 calories, an amount which he found too large to be eaten with comfort, while the later investigator raised the fuel value of his daily food from 2930 to only 3250 calories. As the experiments by Dapper contain other points of interest bearing on the question before us, we may advantageously consider them somewhat in detail. The following table gives the more important results:
No. of Exp. |
Duration. |
Character of Food. |
Food Composition. |
Nitrogen Balance. |
Maxima and Minima of Nitrogen-gain. |
|
|---|---|---|---|---|---|---|
| Nitrogen. | Calories. |
|||||
days |
grams |
grams |
grams |
|||
1 |
6 |
Ordinary mixed diet |
20.25 |
2930 |
+2.18 |
+3.2 on 4th day. +1.5 on 6th day. |
2 |
12 |
Ditto + 80 grams starch |
20.09 |
3250 |
+3.32 |
+4.75 on 2d day. +4.65 on 12th day. +2.30 on 8th day. |
3 |
9 |
Ditto + 80 grams starch, + 40 grams plasmon |
24.58 |
3400 |
+2.55 |
+5.98 on 1st day. +4.73 on 2d day. +0.50 on 6th day. +1.60 on 9th day. |
As we look at these results, the nitrogen gain for the first and second days of the third experiment and the first day of the second experiment may well attract our attention, since they show an astonishing laying by of proteid, or gain of flesh, under the influence of a comparatively small increase in the fuel value of the food. A gain of 5.98 grams of nitrogen means 37.3 grams of proteid, or more than an ounce; by no means an inconsiderable addition for one day to the store of tissue proteid. In the third experiment, where plasmon (dried,101 milk proteid) was added to the diet, there is to be noted a gradual falling off in the proteid-sparing power, which may perhaps be interpreted as implying that the body was practically saturated with proteid, and that owing to this fact the body was unable to continue its laying hold of nitrogen. In the entire period of 21 days, however, the body had succeeded in accumulating a store of 62.8 grams of nitrogen, or 392 grams of proteid, and this without adding very largely to the intake of non-nitrogenous matter. This experiment affords a striking illustration of the ability of the body to “fatten on nitrogen,” but it is very doubtful if such results can generally be obtained. Lüthje,32 however, has reported a large retention of nitrogen on a diet containing 50 grams of nitrogen daily, with a fuel value of 4000 calories. It is more than probable that there existed in these particular cases some personal peculiarity or idiosyncrasy which favored the proteid-sparing power. The personal coefficient of nutrition is not to be ignored; it shows itself in many ways, and the above results are to be counted among those that are exceptional and not the rule. In the words of Magnus-Levy, “a given diet with Cassius may lead to different results than with Anthony.”
For the study of many questions in nutrition, it becomes necessary to determine accurately the transformations of energy within the body as contrasted with the transformation of matter; the total income and outgo of energy, measured in terms of heat, are to be compared one with the other and a balance struck. Further, in studying the metabolism of carbohydrate and fat it is necessary to determine the output of gaseous products through the lungs and skin; to estimate the excretion of carbon dioxide and water, and the intake of oxygen. For these purposes, a special form of apparatus known as a respiration calorimeter is employed. The double102 name is indicative of the twofold character of the apparatus, viz., a suitably constructed chamber so arranged as to permit of measuring at the same time the respiratory products and the energy given off from the body. The form of apparatus best known to-day, and with which exceedingly satisfactory work has been done, is the Atwater-Rosa apparatus, as modified by Benedict. It consists essentially of a respiration chamber, in reality an air-tight, constant-temperature room (with walls of sheet metal, outside of which are two concentric coverings of wood completely surrounding it, with generous air spaces between), sufficiently large to admit of a man living in it for a week or more at a time. Connected with the chamber is a great variety of complex apparatus for maintaining and analyzing the supply of oxygen, determining the amount of carbon dioxide and of water, etc., etc. As an apparatus for measuring heat, the chamber may be described as “a constant-temperature, continuous-flow water calorimeter, so devised and manipulated that gain or loss of heat through the walls of the chamber is prevented, and the heat generated within the chamber cannot escape in any other way than that provided for carrying it away and measuring it.”33
In illustration of the efficiency of an apparatus of this description, and of the close agreement obtainable by direct calorimetric measurement with the estimated energy, as figured from the materials oxidized in the body, we may quote the following data from Dr. Benedict’s report, referred to in the footnote. The subject was a young man who had been fasting for five days. The experiment deals with the metabolism on the first day after the fast, when a diet composed mainly of milk was103 made use of, containing 53.31 grams of proteid, 211.87 grams of fat, and 75.41 grams of carbohydrate. The following table shows the results of the experiment:
Heat of Combustion of Food and Excreta as Determined by Bomb Calorimeter. |
(d) Available Energy from Food. a-(b+c) |
(f) Estimated Energy from Material Oxidized in the Body. d-e. |
Heat Measured by Respiration Calorimeter. |
Heat Measured Greater or Less than Estimated. |
||||
|---|---|---|---|---|---|---|---|---|
(a) Food. |
(b) Excre- ment. |
(c) Urine. |
Amount. |
Propor- tion. |
||||
calories |
calories |
calories |
calories |
calories |
calories |
calories |
calories |
per cent |
2569 |
149 |
103 |
2317 |
+229 |
2088 |
2113 |
+25 |
+1.2 |
As is seen from the above figures, the total fuel value of the food was 2569 calories. The fuel value of the unoxidized portion of the food contained in the excreta was 149 + 103 calories, leaving as the available energy of the food 2317 calories. This must be further corrected by the fact, mentioned in the footnote, that a portion of the food was stored as fat and glycogen, while the body lost at the same time a small amount of proteid. Making the necessary correction for these causes, we find 2088 calories as the energy from material oxidized in the body. The actual output of energy as measured by the calorimeter was 2113 calories, only 1.2 per cent greater than the estimated amount.
By aid of the respiration calorimeter, many important questions in nutrition can be more or less accurately answered, especially such as relate to the total energy requirements of the body. The law of the conservation of energy obtains in the human body as elsewhere, and if we can measure with accuracy the total heat output, with any energy liberated in the form of work, and at the same time determine the total excretion of carbon dioxide, water, nitrogen, etc., together104 with the intake of oxygen, it becomes not only possible to ascertain the energy requirements of the body under different conditions, but, aided by data obtainable through study of the exchange of matter, we can draw important conclusions concerning the sources of the energy, i. e., whether from proteid, fat, or carbohydrate.
It is obvious that a man asleep, or lying quietly at rest, in the calorimeter, especially when he has been without food for some hours, furnishes suitable conditions for ascertaining the minimal energy requirements of the body. Under such conditions, bodily activity and heat output are at their lowest, and we are thus afforded the means of determining what is frequently called the basal energy exchange of the body. The following table taken from Magnus-Levy, and embodying results from many sources, shows the heat production during sleep, calculated for 24 hours, of various individuals of different body-weight and of different body surface.
I venture to present these individual results, rather than make a general statement simply, because it is important to recognize the fact that the basal energy exchange differs according to body-weight, extent of body surface, and the condition of the body. In the table, the results are arranged in the order of body-weight, and it is plain to see that the absolute energy exchange is greater with heavy persons than with light, yet the energy exchange does not increase in proportion to increase of body-weight. With a man of 83 kilos body-weight, the basal exchange is only 30–40 per cent higher than in a man of 43 kilos body-weight. In other words, the man of small body-weight has, per kilo, a much higher basal exchange than the heavier man. The energy exchange is more closely proportional to the extent of body surface than to weight.
105
Body-weight of the Individual. |
Total Calories for 24 Hours. |
Calories per Kilo of Body-weight. |
Body-weight of the Individual. |
Total Calories for 24 Hours. |
Calories per Kilo of Body-weight. |
|---|---|---|---|---|---|
kilos |
kilos |
||||
43.2 |
1333 |
30.9 |
67.5 |
1608 |
23.8 |
48.0 |
1214 |
25.3 |
67.5 |
1621 |
24.0 |
50.0 |
1315 |
25.9 |
70.0 |
1661 |
23.7 |
53.0 |
1527 |
28.8 |
70.0 |
1620 |
23.1 |
55.0 |
1590 |
28.9 |
71.2 |
1787 |
25.1 |
56.5 |
1519 |
26.8 |
72.6 |
1550 |
21.3 |
57.2 |
1560 |
27.3 |
72.7 |
1657 |
22.8 |
58.0 |
1510 |
26.0 |
73.0 |
1584 |
21.7 |
62.5 |
1431 |
22.9 |
73.0 |
1630 |
22.4 |
63.0 |
1418 |
22.5 |
75.6 |
1670 |
22.1 |
63.0 |
1492 |
23.7 |
82.0 |
1556 |
19.0 |
64.0 |
1656? |
25.8 |
82.7 |
2030? |
24.5 |
64.9 |
1475 |
22.7 |
83.5 |
1670 |
20.0 |
65.0 |
1498 |
23.0 |
88.3 |
2019? |
22.9 |
65.0 |
1445 |
22.2 |
90.4 |
1773 |
19.6 |
As Richet has expressed it, the basal energy exchange is inversely proportional to the body-weight and directly proportional to the body surface. This is in harmony with the view advanced by v. Hösslin, “that all the important physiological activities of the body, including of course its internal work and the consequent heat production, are substantially proportional to the two-thirds power of its volume, and that since the external surface bears the same ratio to the volume, a proportionality necessarily exists between heat production and surface.”35
There are, however, many circumstances that modify, or influence, energy exchange. Thus, the taking of food, with all the attendant processes of digestion, assimilation, etc., involves106 an expenditure of energy not inconsiderable. This has been experimentally demonstrated on man by several investigators. With fatty food, Magnus-Levy found that his subject lying upon a couch, as completely at rest as possible, produced in the 24 hours 1547 calories when 94 grams of fat were eaten, and 1582 calories when 195 grams of fat were consumed. The increase of heat production over the basal energy exchange was 10 and 58 calories respectively. With a mixed diet, where proteid food is a conspicuous element, the increase in heat production is much more marked. Thus, in some experiments reported from Sweden the following data were obtained:36
| Day. | Energy of the Food. |
Heat Production. |
|---|---|---|
calories |
calories |
|
| First | 4141 |
. . . . |
| Second | 4277 |
2705 |
| Third | 0 |
2220 |
| Fourth | 0 |
2102 |
| Fifth | 0 |
2024 |
| Sixth | 0 |
1992 |
| Seventh | 0 |
1970 |
| Eighth | 4355 |
2436 |
| Ninth | 3946 |
2410 |
We see here an increase of 495 calories per day in heat production, due to metabolism of the food ingested. In other words, with a basal energy exchange of 2022 calories, the average of the five fasting days, energy equivalent to 495 calories was expended in taking care of the ingested food. It should be added, however, that the daily ration here was107 somewhat excessive, 4193 calories being considerably in excess of the requirements of the body. Finally, it should be stated that of the several classes of foods, proteids cause the greatest increase in metabolism and fats the least.
In studying heat production in the body under varying conditions, one of the important aids in drawing conclusions as to the character of the body material burned up is the respiratory quotient. This is the relationship, or ratio, of the oxygen absorbed to the oxygen of the carbon dioxide eliminated, viz., CO2O2. Carbohydrates (C6H12O6, C12H22O11) all contain hydrogen and oxygen in the proportion to form water, H2O, and in their oxidation they need of oxygen only such quantity as will suffice to oxidize the carbon (C) of the sugar to carbon dioxide (CO2). Carbohydrates, starch and sugars, have a respiratory quotient of 1.00. Fat, on the other hand, has a respiratory quotient of 0.7, and proteid, 0.8. Hence, it is easy to see that the respiratory quotient will approach nearer to unity as the quantity of carbohydrate burned in the body is increased. Similarly, the respiratory quotient will grow smaller the larger the amount of fat burned up. Practically, we never find a respiratory quotient of 1.0 or 0.7, because there is always some oxidation of proteid in the body. If, by way of illustration, we assume that the energy of the body under given conditions comes from proteid to the extent of 15 per cent, while the remaining 85 per cent is derived from the oxidation of carbohydrate, the respiratory quotient will be 0.971. If, however, the 85 per cent of energy comes from fat, the respiratory quotient will change to 0.722. In the resting body, as in the early morning hours, after a night’s sleep and before food is taken, the respiratory quotient is generally in the neighborhood of 0.8. When, however, as sometimes happens, the quotient at this time of day approaches 0.9, it must be assumed that sugar is108 being burned in the body, presumably from carbohydrate still circulating from the previous day’s intake.
As can easily be seen, any special drain upon either fat or carbohydrate in the processes of the body will be indicated at once by a corresponding change in the respiratory quotient. This we shall have occasion to notice later on, in considering the source of the energy of muscle contraction. Further, the respiratory quotient will naturally change in harmony with transformations in the body which involve alterations in oxygen-content, without the oxygen of the inspired air being necessarily involved; as in the formation of a substance poor in oxygen, such as fat, from a substance rich in oxygen, such as carbohydrate. Moreover, the reversal of this reaction, as in the formation of sugar from proteid with a taking on of oxygen, will produce a corresponding effect upon the respiratory quotient. As Magnus-Levy has clearly pointed out, in the formation of fat from carbohydrate, carbon dioxide is produced in large amount without the oxygen of the inspired air being involved at all. In such a change, 100 grams of starch will yield about 42 grams of fat, while at the same time 45 grams of carbon dioxide will be produced. This might cause the respiratory quotient to rise as high as 1.38. Again, in the formation of sugar from proteid, the respiratory quotient may sink very decidedly, the changes involved being accompanied by a taking on of oxygen from the air, without, however, any corresponding increase of carbon dioxide in the expired air. Assuming a manufacture of 60 grams of dextrose from 100 grams of proteid, i. e., from the non-nitrogenous moiety of the proteid molecule, a respiratory quotient of 0.613 would be possible. Thus, a diabetic patient, living upon a carbohydrate-free diet, consuming only proteid and fat, may show a respiratory quotient of 0.613–0.707. These illustrations will suffice to show how chemical alterations taking place in the body, involving transformations of109 proteid, fat, and carbohydrate of the tissues and of the food, may produce alterations in the respiratory quotient without necessarily being directly connected with intake of oxygen or output of carbon dioxide through the lungs; and how, conversely, the respiratory quotient becomes a factor of great significance in throwing light upon the character of the nutritive changes taking place in the body.
Among the various conditions that influence the energy exchange of the body, muscle work stands out as the most conspicuous. It needs no argument to convince one that all forms of muscular activity involve liberation of the energy stored up in the tissues of the body; and consequently that all work accomplished means chemical decomposition, in which complex molecules are broken down into simple ones with liberation of the contained energy, the energy exchange being proportional to the amount of work done. As we have seen, the basal energy exchange of the normal individual is ascertained by studying his heat production while at rest—best during sleep—without food, when involuntary muscle activity and heat production are at their lowest. The maximum energy exchange is seen in the individual at hard muscular work. Heat production is then at its highest, as can be ascertained by direct calorimetric observation; or, by studying the output of excretory products, which measure the extent of the oxidative processes from which comes the energy for the accomplishment of the work. As an illustration of the general effect of muscular work on the energy exchange of the body, we may cite a summary of some results reported by Atwater and Benedict,37 the figures given being average results, from several individuals, and covering different periods of time. Though not strictly comparable in all details, they are sufficiently so to illustrate the main principle.
110
HEAT GIVEN OFF BY BODY, INCLUDING FOR WORK EXPERIMENTS THE HEAT EQUIVALENT OF THE EXTERNAL MUSCULAR WORK.
Kind of Experiment. |
Total Amount of Heat in 24 Hours. |
Rates per Hour. |
Average for 24 Hours. |
||||
|---|---|---|---|---|---|---|---|
Day Periods. |
Night Periods. |
||||||
7 A. M. to 1 P. M. |
1 P. M. to 7 P. M. |
7 P. M. to 1 A. M. |
1 A. M. to 7 A. M. |
||||
calories |
calories |
calories |
calories |
calories |
calories |
||
Rest experiments |
2262 |
106.3 |
104.4 |
98.3 |
67.9 |
94.3 |
|
Work experiments Heat eliminated |
4225 |
231.7 |
235.6 |
118.1 |
78.4 |
166.6 |
|
Heat equivalent of external muscular work |
451 |
58.5 |
56.8 |
. . . |
. . . |
. . . |
|
Total |
4676 |
290.2 |
292.4 |
118.1 |
78.4 |
194.8 |
|
The work done in these experiments was on a stationary bicycle in the calorimeter, and the heat equivalent was calculated from measurements made by an ergometer attached to the bicycle. We are not concerned here with details, but simply with the general question of the influence of muscular work upon the energy exchange of the body. We note that the work of the day periods, 7 A. M. to 7 P. M., resulted, in the several cases brought together under the average figures, in an increased heat production amounting to more than 100 per cent. Further, we observe that in the body, as in all machines, only a fraction of the energy liberated by the accelerated chemical decomposition, or oxidation, was manifested as mechanical work, the larger part by far being heat eliminated and lost. Thus, Zuntz has found that, in man, about 35 per cent of the extra energy of the food used in connection with external muscular work is available for that111 work. This, however, shows a noticeably higher degree of efficiency than is generally obtainable by the best steam or oil engines. Lastly, attention may be called to the fact that after the work of the day was finished at 7 P. M., the next period of six hours still showed an accelerated metabolism, as contrasted with what took place during absolute rest.
As bearing upon the exchange of matter in the body in connection with muscular work, and as showing the relationship which exists here between energy exchange and exchange of matter, we may quote a few data relating to the elimination of carbon dioxide; remembering that this substance represents particularly the final oxidation product in the body of carbonaceous materials, such as fat and carbohydrate. The following data, taken from Atwater and Benedict,38 being results of experiments upon the subject “J. C. W.,” are of value as showing the variations in output of carbon dioxide that may be expected under the conditions described:
Period. |
Rest Experiments without Food. |
Rest Experiments with Food. |
Work Experiments with Diet. |
Work Experiments with Fat Diet. |
Extra Se- vere Work Experiment with Fat Diet. |
|---|---|---|---|---|---|
grams |
grams |
grams |
grams |
grams |
|
| 7 A. M. to 1 P. M. | 189.6 |
230.4 |
694.0 |
642.3 |
907.0 |
| 1 P. M. to 7 P. M. | 172.6 |
232.0 |
705.6 |
634.8 |
821.3 |
| 7 P. M. to 1 A. M. | 167.2 |
196.6 |
260.1 |
230.3 |
842.7 |
| 1 A. M. to 7 A. M. | 146.7 |
153.1 |
161.1 |
157.6 |
502.6 |
| Total for 24 hours | 676.1 |
812.1 |
1820.8 |
1665.0 |
3073.6 |
In considering these figures bearing on the output of carbon dioxide under the conditions specified, we note at once a correspondence with the total energy exchange, as indicated112 in the preceding table. As previously stated, we are at present dealing simply with generalities, and the important point to be observed here is that muscular work—7 A. M. to 7 P. M.—in the work experiments, increases enormously the output of carbon dioxide. We see clearly emphasized a connection between the total energy exchange of the body, as expressed in calories or heat units, and the oxidation of carbonaceous material, of which carbon dioxide is the natural oxidation product. We note that on the cessation of work—7 P. M. to 7 A. M.—the output of carbon dioxide tends to drop back to the level characteristic of the corresponding period in rest, with or without food. In the experiment with “extra severe muscular work,” the results are different simply because here the subject worked sixteen hours, necessitating a portion of the work being done at night-time. Finally, it should be mentioned that the differences in output of carbon dioxide in these experiments are somewhat greater than in many experiments of this type, although all show the same general characteristics. This may be explained, as stated by the authors from whom the data are taken, “by the fact that J. C. W. was a larger and heavier man than any of the others; that the differences in diet were wider, and that the amounts of external muscular work were larger in these experiments than in those with the other subjects.”
If we pass from experiments of this type, conducted in a calorimeter, to those cases where competitive trials of endurance are held by trained athletes, i. e., where external muscular activity is pushed to the extreme limit, we then see even more strikingly displayed the effect of work in increasing the energy exchange of the body. One of the best illustrations of this type of experiment is to be found in the observations made in connection with the six-day bicycle race held in New York City, at the Madison Square Garden, in113 December, 1898.39 The observations in question were made upon three of the athletes, one of whom withdrew early in the fourth day, while the others continued until the close of the race—142 consecutive hours—winning the first and fourth places, respectively. The following table gives the computation of energy of the material metabolized, exclusive of body-fat lost:
Subject. |
Duration of Experiment. |
Total Energy Metabolized. |
Average per Day. |
|---|---|---|---|
days |
calories |
calories |
|
| Miller | 6 |
28917 |
4820 |
| Albert | 6 |
36441 |
6074 |
| Pilkington | 3 |
13301 |
4464 |
Miller, the winner of the race, who averaged a daily energy exchange of 4820 calories, rode 2007 miles during the week, and finished the race without physical or mental weakness resulting from the fatigue and strain. During the first five days, he rode about 21 hours a day and slept only 1 hour. Albert, who weighed a few pounds less than Miller, covered 1822 miles in 109 hours, with an average daily exchange of 6074 calories. We may add a table (on the following page) showing the balance of income and outgo of nitrogen in these three subjects, as being of general interest in this connection. The figures given are averages per day.
Subject. |
Duration of Exp. |
Income in Food. |
Nitrogen. |
||||||
|---|---|---|---|---|---|---|---|---|---|
Proteid. |
Fat. |
Carbo- hydrate. |
Fuel Value. |
In Food. |
In Urine. |
In Excre- ment. |
Loss. |
||
days |
grams |
grams |
grams |
calories |
grams |
grams |
grams |
grams |
|
| Miller | 6 |
169 |
181 |
585 |
4770 |
29.4 |
36.2 |
1.8 |
8.6 |
| Albert | 6 |
179 |
198 |
559 |
6095 |
29.1 |
33.7 |
2.5 |
7.1 |
| Pilkington | 3 |
211 |
178 |
509 |
4610 |
36.0 |
38.9 |
2.2 |
5.1 |
The special significance of these data, as bearing upon the topic under discussion, is that apparently all three of the subjects were drawing in a measure upon their body material. As stated by Atwater and Sherman, Pilkington lost per day 5.1 grams of nitrogen; that is to say, the total nitrogen excreted114 exceeded the total nitrogen of the food by 5.1 grams per day, corresponding to 33 grams of proteid, which must have been drawn from the supply in the body. If we assume that lean flesh contains 25 per cent of proteid, this would mean about 4 34 ounces per day. The other two subjects, Miller and Albert, lost from the body per day 8.6 grams and 7.1 grams respectively of nitrogen, which would imply a loss of about 54 grams and 44 grams of body proteid respectively, or 8 ounces and 6 14 ounces of lean flesh per day. It is evident, therefore, that none of the three subjects consumed sufficient food to avoid loss of body proteid, under the existing conditions of muscular activity. Indeed, it may be noted in Miller’s case that the average fuel value of the food per day was 4770 calories, while the average expenditure of energy per day was 4820 calories. We should naturally expect, however, that any small deficiency in fuel value would be made good by a call upon body fat. “Why the body should use its own substance under such circumstances is a question which at present cannot be satisfactorily answered. The fact that such was the case, each of the contestants who finished the race consuming during the period body protein equivalent to 2 or 3 pounds of lean flesh, and that no injury resulted therefrom, would seem to indicate that these men had stores of protein which could be metabolized115 to aid in meeting the demands put upon the body by the severe exertion, without robbing any of the working parts, and at the same time relieving the system of a part of the labor of digestion. Possibly, the ability to carry such a store of available protein is one of the factors which make for physical endurance.”40 This possibility we shall have occasion to discuss in another connection. At present, the facts presented are to be accepted as accentuating the general law that the energy exchange of the body, everything else being equal, is increased proportionally to increase in the extent of external muscular activity. It may be noted that Albert, who did considerably less work than Miller, showed a much larger exchange of energy than the latter athlete. This, however, is to be connected with the fact that his fuel intake was 1300 calories larger per day than Miller’s; in other words, the conditions were not equal. This fact also calls to mind the observations of Schnyder,41 who, studying the relationship between muscular activity and the production of carbon dioxide, maintained that the quantity of this excretory product formed depends less upon the amount of work accomplished than upon the intensity of the exertion; efficiency in muscular work varying greatly with the condition of the subject, and his familiarity with the particular task involved.
From what has been said, it is obvious that oxygen consumption, as well as output of carbon dioxide, must vary enormously with variations in the muscular activity of the body. The one important factor influencing the quantities of oxygen and carbon dioxide exchanged in the lungs, i. e., the extent of the respiratory interchange, is muscular activity; and since, as we have seen, carbonaceous material is the substance mainly oxidized in muscle work, it follows, as carbon116 dioxide is excreted principally through the lungs, that the respiratory interchange becomes in good measure an indicator of the extent of chemical decomposition incidental to external work. If we recall that man, on an average, at each inspiration draws in about 500 cubic centimeters of air (30 cubic inches), and that for the 24 hours he averages 15 breaths a minute, it is easy to see that in one minute the average man will inspire 7.5 litres of air, or 450 litres an hour, with a total of 10,800 litres for the entire day, which is equivalent to about 380 cubic feet. This would be a volume of air just filling a room 7 13 feet in length, width, and height. Inspired air loses to the body 4.78 volumes per cent of oxygen, while expired air contains an excess of 4.34 volumes per cent of carbon dioxide. In muscular work, respiration is increased in frequency and in depth. The volume of air exchanged in the lungs during severe labor may be increased sevenfold, while oxygen consumption and carbon dioxide excretion are frequently increased 7–10 times. The following figures, being values for one minute, show the effect on oxygen consumption of walking on a level and climbing, the subject being a man of 55.5 kilos body-weight:42
| Form of Work. | Oxygen Consumption in Cubic Centimeters. |
Respiratory Quotient. |
||
|---|---|---|---|---|
Total. |
After Deducting Value for Rest. |
|||
Total. |
For Each Kilo of Moving Weight. |
|||
| Standing at rest | 263.75 |
. . . . |
. . . . |
0.801 |
| Walking on a level | 763.00 |
499.25 |
8.990 |
0.805 |
| Climbing | 1253.20 |
989.45 |
17.819 |
0.801 |
Remembering that these figures represent the oxygen consumption for only one minute of time, it is easy to see the striking effect of moderate and vigorous exercise on respiratory interchange. Simply walking along a level suffices to increase the consumption of oxygen threefold over what occurs when the body stands at rest. When the more vigorous exercise attendant on lifting the body up a steep incline is attempted, most striking is the great increase in the amount of oxygen consumed. We thus see another forcible illustration of the influence of muscular activity upon the exchange of matter in the body, and a further confirmation of the statement, so many times made, that oxidation—especially the oxidation of fats and carbohydrates by which large quantities of heat are set free, easily convertible into mechanical energy—is a primary factor in the metabolic processes, by which the machinery of the living man is able to work so efficiently.
Finally, we cannot avoid the conclusion that the outgoings of the body, in the form of matter and energy, are subject to great variation, incidental to the degree of activity of the day or hour. The ordinary vicissitudes of life, bringing days of physical inaction, followed perhaps by periods of unusual activity; changes in climatic conditions, with their influence upon heat production in the body; alterations in the character and amount of the daily dietary, etc.,—all seemingly combine as natural obstacles to the maintenance of a true nutritive balance. Outgo, however, must be met by adequate amounts of proper intake if there is to be an approach toward a balance of nutrition. In some way the normal, healthy man does maintain, approximately at least, a condition of balance; not necessarily for every hour or for every day, but the intake and outgo if measured for a definite period, not too short, say for a week or two, will be found to approach each other very closely. Body equilibrium and118 approximate nitrogen balance may be reasonably looked for, as well as a balance of total energy, in the case of a healthy man leading a life which conforms to ordinary physiological requirements. The man who, on the other hand, consciously or unconsciously, continues an intake way beyond the outgo, whose daily income of nitrogen and total fuel value far exceeds the requirements of his body, obviously lives with an accumulating plus balance, which ordinarily shows itself in increasing body-weight and with a storing away of fat.
Equally conspicuous is the effect of an inadequate income of proper nutriment; a food supply which persistently fails to furnish the available nitrogen and total energy value called for by the body under the conditions prevailing, will inevitably result in a minus balance, which, if continued too long, must of necessity tax the body’s store to the danger limit. At the same time, the well-nourished individual, without being unduly burdened by a bulky store of energy-containing material, is always supplied with a sufficient surplus to meet all rational demands, when from any cause the intake fails, for brief periods of time, to be commensurate with the needs of the body. It is reasonable to believe, however, that in the maintenance of good health, and the preservation of a high degree of efficiency, the body should be kept in a condition approaching a true nutritive balance.
119
SOURCE OF THE ENERGY OF MUSCLE WORK, WITH SOME THEORIES OF PROTEID METABOLISM
Topics: Relation of muscle work to energy exchange. Views of Liebig. Experimental evidence. Relation of nitrogen excretion to muscle work. Significance of the respiratory quotient in determining nature of the material oxidized. Fats and carbohydrates as source of energy by muscles. Utilization of proteid as a source of energy. Formation of carbohydrate from proteid. Significance of proteid metabolism. Theories of Carl Voit. Morphotic proteid. Circulating proteid. General conception of proteid metabolism on the basis of Voit’s theories. Pflüger’s views of proteid metabolism. Rapidity of elimination of food nitrogen. Methods by which nitrogen is split off from proteid. Theories of Folin. Significance of creatinin and of the percentage distribution of excreted nitrogen. Endogenous or tissue metabolism. Exogenous or intermediate metabolism. Needs of the body for proteid food possibly satisfied by quantity sufficient to meet the demands of tissue or endogenous metabolism. Bearings of Folin’s views on current theories and general facts of proteid metabolism. Large proteid reserve and voluminous exogenous metabolism probably not needed. Importance of feeding experiments in determining the true value of different views.
As we have already seen, every form of muscular activity begets an increase in the energy exchange of the body. Between the two extremes of absolute rest and excessive muscular exertion, we find differences of 2000 calories or more per day as representing the degree of chemical decomposition corresponding to the particular state of muscular activity. The work of the involuntary muscles, such as have to do with peristalsis, respiration, rhythmical beat of the heart, etc., is a relatively constant factor, though naturally subject to some variation, as has been pointed out in other120 connections. External muscular activity, however, is the one factor above all others that modifies the rate of energy exchange. A little longer walk, a heavier load to carry, a steeper hill to climb, any increase great or small in the activity of the muscles of the body, means a corresponding increase in chemical decomposition, with increased output of the ordinary products of tissue oxidation. The material so consumed, or oxidized, must be made good to hold the body in equilibrium; the supplies drawn upon are to be replaced, if the tissues of the body are to be kept in a proper state of efficiency.
What is the nature of the material used up in connection with muscle work? As can readily be seen, this is an important question, for on its answer depends, in some measure at least, the character of the proper intake, or food, to be supplied in order to make good the loss. If the energy of mechanical work, the energy of muscle contraction, comes from the breaking down of proteid matter alone, then obviously excessive muscular work would need to be accompanied, or followed, by a generous supply of proteid food. If, on the other hand, external work means liberation of energy solely from non-nitrogenous materials, then it is equally clear that fats and carbohydrates are the proper foods to offset the drain incidental to vigorous muscular action.
The views of Liebig, briefly referred to in a previous chapter, held sway over physiologists for many years. His dictum that proteid foods were true plastic foods, entering into the structure of the tissues of the body, and that they alone were the real sources of muscular energy, met for a time with no opposition. It was not until the advent of a more critical spirit, accompanied by a fuller appreciation of the necessity of experimental evidence, that physiologists began to test with scientific accuracy the validity of the current views. It is worthy of note that long prior to this time, even before oxygen was discovered, the far-sighted and resourceful121 John Mayow, in his work with the various “spirits” of the body and their relation to respiration, etc., evolved the view that muscular power has its origin in the combustion of fat brought to the muscles by the blood and burned there by aid of a gas or “spirit” taken from the air by the lungs, and likewise carried to the muscles by the circulating blood. Considering the time when Mayow lived and the dearth of true scientific knowledge as we measure it to-day, his hypothesis was a wonderful forestalling of present views.
It is quite obvious that the views of Liebig, if true, admit of easy proof; since, if the energy of muscular power comes from the breaking down of proteid, there should be a certain parallelism between the output of nitrogen from the body and the amount of muscular work accomplished, everything else being equal. As stated in a previous chapter, such study of this question as was made soon disclosed the fact that the one element above all others that seemed to influence the output of nitrogen was the intake of proteid food. Thus, the English investigators, Lawes and Gilbert, found by experimenting with animals that when the latter were kept under uniform conditions of muscular work, the amount of nitrogen excreted ran parallel with the intake of nitrogen. Further, in the early experiments of Voit, the results obtained clearly showed that variations in the amount of work performed were practically without influence on the excretion of nitrogenous waste products.
The experiment, however, that came as a death blow to the theories of Liebig was that of Fick and Wislicenus,43 who in 1865 made an ascent of the Faulhorn, 6500 feet high, using a diet wholly non-nitrogenous. From the nitrogen excreted they were able, of course, to calculate the amount of proteid oxidized in the body during the period122 of work, and found that the proteid consumed could not have furnished, at the most, more than one-half the energy required to lift the weights of their bodies to the top of the high peak. Further, they observed that neither during the work period, nor immediately after, was there any noticeable increase in the excretion of nitrogen. Obviously, as they state, the oxidation of proteid matter in the body cannot be the exclusive source of the energy of muscular contraction, since the measurable amount of external work performed in the ascent of the mountain was far greater than the equivalent of the energy capable of being furnished by the proteid actually burned. To which may be added the fact that considerable energy, not measurable in their experiment, must have been employed in the work of the involuntary muscles of the body; thus increasing by so much the difference between the muscular work actually accomplished and the available energy from proteid consumed. It is true that minor criticisms regarding certain details of the experiment can be offered to-day, such as the fact that the men were, in a measure, in a state of “nitrogen starvation,” etc., but these criticisms do not in any degree militate against the main thesis that the energy of muscular contraction does not come exclusively from the consumption or breaking down of proteid, either of food or tissue. Vigorous and even severe muscular work does not necessarily increase the decomposition of proteid material. Dogs made to run in large treadmills, with the same diet as on resting days, were found to excrete practically no more nitrogen than during the days of rest. Occasionally, however, in some one experiment the output of nitrogen would show an increase over the output on resting days. Further, experiments made with horses led to essentially the same result, except that greater increase in the excretion of nitrogen was observed than with dogs. This increase in nitrogen output, however, as a concomitant123 of increased muscular activity, could be prevented by adding to the amount of carbohydrate food.
While experiments of this nature, on man and animals, all tended to show little or no increase in the excretion of nitrogen, as a result of muscle work; and likewise no increase in the output of sulphur and phosphorus, thus strengthening the view that muscular energy is not the result of proteid disintegration, there was observed marked increase in the consumption of oxygen, and in the excretion of carbon dioxide. Non-nitrogenous matter was thus at once suggested as the material with which muscle chiefly does its work. There is to-day no question of the general truth of this statement, yet there are other aspects of the problem to be considered before we can lay it aside. Pflüger, working with dogs, and Argutinsky, experimenting on himself by arduous mountain climbing, reached conclusions seemingly quite opposed to what has just been said. Their results, however, admit of quite a different interpretation from what they were disposed to attach to them. Thus, Pflüger44 would go back to the old view that all muscle work is at the expense of proteid material, because lean dogs fed mainly, or entirely, on meat and made to do an excessive amount of work were found by him to excrete nitrogen somewhat in proportion to the amount of work done. Argutinsky,45 likewise, in his mountain climbing carried to the point of fatigue, and with a high proteid intake likewise, saw in the increased output of nitrogen a suggestion of the same idea. In reality, however, their results merely prove that, under some circumstances, proteid may serve as the chief source of muscular energy; as when the body is poor in fat and carbohydrate, or when the intake consists solely of proteid matter. In other words, muscular work may result in124 an increased excretion of nitrogen when the work is very severe, and there is not a corresponding increase in the fats or carbohydrates (fuel ingredients) of the food. In the words of Bunge,46 “we might assume à priori, on teleological grounds, that in the performance of its most important functions the organism is to a certain extent independent of the quality of its food. As long as non-nitrogenous food is supplied in adequate quantity or is stored up in the tissues, muscular work is chiefly maintained from this store. When it is gone the proteids are attacked.”
There is no question that the energy of muscular contraction can come from all three classes of organic foodstuffs. Voluntary muscular movement is under the control of the nervous system, and when the stimulus is applied the muscle is bound to contract, provided of course there is sufficient energy-containing material present to furnish the means. Muscle tissue, like other tissues and organs, has a certain power of adaptability, by which it is able to do its work, even though it is not adequately supplied with its preferred nutrient. While proteid is plainly not the material from which the energy of muscular contraction is ordinarily derived, it is equally evident that in emergency, as when the usual store of carbohydrate and fat is wanting, proteid can be drawn upon, and in such cases vigorous work may be attended with increased nitrogen output. In harmony with this statement, we find on record in recent years many experiments, both with man and animals, where severe muscular labor is accompanied by an excretion of nitrogen beyond what occurs on days of rest; but by simply adding to the intake of non-nitrogenous food this increased outgo of nitrogen is at once checked. With moderate work, the nitrogen outgo is rarely influenced; it is only when the work becomes125 excessive, or the store of non-nitrogenous reserve is small and the intake of the latter food is limited, that proteid matter is drawn upon to supply the required energy.
Recalling what has been said regarding the significance of the respiratory quotient, it is obvious that we have here a means of acquiring information as to the character of the material that is burned up in the body during muscular work. Increased metabolism of carbohydrate will necessarily result in raising the respiratory quotient, and if the latter food material alone is involved the respiratory quotient must naturally approach 1.0. Zuntz, however, has clearly shown that vigorous muscular activity does not materially change the respiratory quotient; except in cases of very severe work, where the oxygen-supply of the muscles is interfered with. Indeed, the muscles may be made to do work sufficient to increase the consumption of oxygen threefold or more, without any change in the respiratory quotient being observed. And as there is frequently no change whatever in the output of nitrogen under these conditions, it follows that the energy of the muscle work must have come from the decomposition of non-nitrogenous material. If carbohydrates alone were involved, the respiratory quotient would obviously undergo change. Since, however, this remains practically stationary, we are led to the conclusion that fat must be involved in large degree, in addition to carbohydrate.
In this connection, it is a significant fact that with fasting animals, where the store of carbohydrate material is more or less used up, severe muscle work may be accomplished without any appreciable increase in nitrogen output, thus showing that proteid material is not involved and clearly pointing to fat as the source of the muscular energy. Thus, in an experiment referred to by Leathes, a dog on the sixth and seventh day of starvation was made to do work in a treadmill equivalent to climbing to a height of 1400 meters, yet the output of126 nitrogen was increased from six to only six and a half grams. Obviously, not much of the energy of this muscle work could have come from the breaking down of proteid, but it must have been derived mainly from the oxidation of fat. There is abundant evidence that fat can be used as a source of energy by muscles, as well as carbohydrates and proteids, and there is every reason for believing that the yield of work for a given amount of chemical energy in the form of fat is as good as in the case of either of the other two substances. In fact, the observations of Zuntz show that fat can be used just as economically by the body for muscle work as either carbohydrates or proteid. Thus, in one experiment,47 he determined the oxygen-consumption and respiratory quotient in a man resting and working on three different diets—one principally fat, one principally carbohydrate, and the other principally proteid—and found that slightly less oxygen and energy were required to do work on the fat diet than on the others. This is clearly shown in the following table:
| Diet Principally. |
Resting. |
Working. |
Kilo- gram- meters of Work Done. |
Per Kilogram-meter of Work. |
|||
|---|---|---|---|---|---|---|---|
| Oxygen Used per Minute. |
Respira- tory Quotient. |
Oxygen Used per Minute. |
Respira- tory Quotient. |
Oxygen Used. |
Calories. |
||
c.c. |
c.c. |
c.c. |
|||||
| Fat | 319 |
0.72 |
1029 |
0.72 |
354 |
2.01 |
9.39 |
| Carbohydrate | 277 |
0.90 |
1029 |
0.90 |
346 |
2.17 |
10.41 |
| Proteid | 306 |
0.80 |
1127 |
0.80 |
345 |
2.38 |
11.35 |
From these data, we see that per kilogram-meter of work less energy was required and less oxygen consumed with fat than with either of the other two foodstuffs; but practically, fat and carbohydrate as sources of muscle energy have about the same value.
127
Much stress is ordinarily laid upon the importance of a large intake of proteid food whenever the body is called upon to perform severe, or long-continued, muscular work; but in view of what has been stated it may be questioned whether there is any real physiological justification for such custom. The pedestrian Weston,48 who in 1884 walked 50 miles a day for 100 consecutive days, was found by Blyth during a period of five days to consume in his food 37.2 grams of nitrogen a day, while he excreted only 35.3 grams, leaving a balance of 1.9 grams of nitrogen per day apparently stored in the body. His daily food during this period was composed of 262 grams of proteid, 64.6 grams of fat, and 799 grams of carbohydrate, with an estimated fuel value of 4850 calories. Yet he performed this large amount of work daily, and still laid by a certain amount of proteid on a ration, the energy value of which would not ordinarily be considered high for the muscular work to be done. Fourteen years prior to this, Weston, while in New York, was carefully studied by Dr. Flint during a period of 15 days, on 5 of which he walked a total of 317 miles. His diet was128 essentially a proteid diet, consisting principally of beef extract, oatmeal gruel, and raw eggs. Nitrogen intake and output were carefully compared during the days of rest and during the days of work, with the results tabulated.
Period. |
Occupation. |
Duration of Test. |
Nitrogen. |
||||
|---|---|---|---|---|---|---|---|
In Food. |
In Urine. |
In Excre- ment. |
Gain + or Loss - |
||||
days |
grams |
grams |
grams |
grams |
|||
| Fore period | Comparative rest | 5 |
22.0 |
18.7 |
1.4 |
+1.9 |
|
| Working period | Walking 62 miles per day |
5 | 13.2 |
21.6 |
1.6 |
–10.0 |
|
| After period | Rest | 5 |
28.6 |
22.0 |
2.2 |
+4.4 |
|
In this case it will be noted that the daily ration was comparatively small, and, further, that during the working period the subject consumed much less proteid than on the resting days. Moreover, when we remember that the total energy value of his diet must have been quite small, it is not at all strange that in the laborious task of walking 62 miles a day he should have temporarily drawn upon his store of body proteid to the extent of 62.5 grams, or 10 grams of nitrogen a day. Such experiences, however, do not by any means constitute proof that in excessive muscular work there is need for the consumption of correspondingly increased quantities of proteid food, or that the energy of muscular work comes preferably from the breaking down of proteid material. Carbohydrate and fat unquestionably take precedence over proteid in this respect, and we may accept as settled the view that in all practical ways carbohydrate and fat stand on an equal footing as sources of muscular energy. Less clear, perhaps, is the question as to how these two radically different types of organic material are utilized by the muscle. It has been a favorite belief among some physiologists that the contracting muscle makes use of only one substance as the direct source of its energy, and that this substance is the sugar dextrose. This view would seemingly imply that fat and proteid must undergo alteration prior to their utilization by the muscle; that, possibly, the carbon of the fat and proteid is transformed into sugar before the muscle can make use of it. So far as fat is concerned, this view is not supported by the facts available, since experiments show that the heat and energy liberated in the utilization of a given amount of fat in muscle work are in harmony with the energy value of the129 fat; in other words, the fat is apparently burned, or oxidized, directly, without undergoing previous transformation into any form of carbohydrate; or, if transformation does occur, under some conditions, it must take place within the muscle and without loss of energy. The practical significance of these facts is at once apparent, for if fat, in order to be available as a source of muscle energy, must first undergo conversion into sugar, it would be far more economical from a physiological standpoint to replace the fat of the diet with carbohydrate in any attempt to provide suitable nourishment for the working muscle. We may safely conclude, however, that fat and carbohydrate, as previously suggested, are in reality both capable of direct metabolism by the muscular tissue, and that each is of value as a source of muscular energy in proportion to its heat of combustion, yielding substantially the same proportion of its potential energy in the form of mechanical work.
Regarding the utilization of proteid as a source of energy by the muscle, there are many grounds for believing that here the body has to deal with certain alterations, before the proteid can be made available. We may indeed conjecture the transformation of a non-nitrogenous portion of the proteid molecule into carbohydrate, as a necessary step in its utilization for muscle work. It is certainly true that in the ordinary katabolic processes, through which proteid passes, there is a tendency for the nitrogen-containing portion to be quickly split off and eliminated, leaving a carbonaceous residue which may represent as much as 80 per cent of the total energy of the original proteid. This so-called carbon moiety of the proteid molecule is apparently much less rapidly oxidized than the nitrogenous portion, and may indeed be temporarily stored in the body, in the form of fat or carbohydrate.13049 We have very convincing proof that the carbohydrate glycogen can be formed from proteid. Thus, the feeding of proteid to warm-blooded animals may be accompanied by an accumulation of glycogen in the liver. This is interpreted as meaning that in the cleavage of proteid by digestion the various nitrogenous products formed are somewhere, probably in the liver, still further acted upon; the contained nitrogen with some of the carbon being converted into urea, while the non-nitrogenous residue is transformed into glycogen, or sugar. That some such change takes place, or, more specifically, that carbohydrate does result from proteid is more strikingly shown in human beings suffering with diabetes. In severe forms of this disease, all carbohydrate food consumed is rapidly eliminated through the kidneys in the form of sugar, the body having lost the power of burning sugar. If such a person is placed upon a diet composed exclusively of proteid, sugar still continues to be excreted, and there is observed a certain definite relationship between the nitrogen output and the excretion of sugar, thus implying that they have a common origin.
Further, there are certain drugs, such as phloridzin, which, when introduced into the circulation, set up a severe diabetes and glycosuria. Dogs treated in this way, fed solely on proteid or even starved for some time, will continue to excrete sugar, and as in the previous instance, there is observed a certain definite ratio between the nitrogen output and the elimination of sugar; thus leading to the conclusion that both arise from the destruction of the proteid molecule. Careful study of this ratio of dextrose to nitrogen has led Lusk to the conclusion that full 58 per cent of the proteid may undergo conversion into sugar in the body. Hence, it is easy to see how in muscle work, when proteid is the sole source of the energy of muscular contraction, the work accomplished may still result from the131 direct oxidation of carbohydrate material, indirectly derived from the proteid molecule. It requires no argument, however, to convince one that such a procedure for the normal individual is less economical physiologically than a direct utilization of carbohydrate and fat, introduced as such and duly incorporated with the muscle substance. Consequently, in the nourishment of the body for vigorous muscular work, there is reason in a diet which shall provide an abundance of carbohydrate and fat; proteid being added thereto only in amounts sufficient to meet the ordinary requirements of the body for nitrogen and to furnish, it may be, proper pabulum for the development of fresh muscle fibres, where, as in training, effort is being made to strengthen the muscle tissue and so enable it to do more work. Increase in proteid food may help to make new tissue, but the source of the energy of muscle work is to be found mainly in the breaking down of the non-nitrogenous materials, carbohydrate and fat.
In view of these facts, we may advantageously consider next the real significance of the proteid metabolism of the body. As we have seen, a meal rich in proteid leads at once—within a few hours—to an excretion of urea equivalent to full 50 per cent of the nitrogen of the ingested proteid, while a few hours later finds practically all of the nitrogen of the intake eliminated from the body. Further, it is to be remembered that in a general way this occurs no matter what the condition of the body may be at the time and no matter how large or small the amount of proteid consumed. In other words, there is practically no appreciable storing of nitrogen or proteid for future needs,—at least none that is proportional to the increase in nitrogen intake, even though the body be in a condition approximating to nitrogen starvation. Moreover, it is to be recalled that the increased proteid metabolism attendant on increased intake of proteid food132 is accompanied by an acceleration of the metabolism of non-nitrogenous matter; thus resulting in a stirring up of tissue change, with consequent oxidation and loss of a certain proportion of accumulated fat and carbohydrate. Coincident with this increased excretion of nitrogen, the output of carbon dioxide is likewise increased somewhat, due as is believed mainly to increased metabolism of the involuntary muscle fibres of the gastro-intestinal tract, by action of which the accelerated peristalsis so characteristic of food intake is accomplished. Further, the increased output of carbon dioxide, under these conditions, is to be attributed also to the greater activity of the digestive and excretory organs, naturally stimulated to greater functional power by the presence of proteid foods and their decomposition products. Still, as stated by Leathes, “the two main end-products of proteid metabolism, urea and carbonic acid, are, to a great extent, produced independently of each other, and the reactions which result in the discharge of the nitrogen are not those in which energy is set free, work done, and carbonic acid produced.” In other words, there is suggested what we have already referred to, viz., that in proteid metabolism a nitrogenous portion of the proteid molecule is quickly split off and gotten rid of, while the non-nitrogenous part may be reserved for future oxidation, serving as a source of muscle energy or for other purposes. This being so, it is plain that “proteid metabolism in so far as it is concerned with the evolution of energy, proteid metabolism in its exothermic stages, may be almost entirely non-nitrogenous metabolism” (Leathes).
Is there any advantage to the body, however, in this carbonaceous residue of the proteid molecule over simple carbohydrate and fat? Can the processes of the body be accomplished more economically, or more advantageously, with a daily diet so constructed that the tissues and organs must133 depend mainly upon this carbon moiety of the proteid molecule for their energy-yielding material? It has been one of the physiological dogmas of the past, that the tissues and organs of the body, or rather their constituent cells, preferred to use proteid for all their needs whenever it was available. If proteid were wanting, either because of insufficient intake, or because of excessive activity, then the tissue cells would draw upon their store of non-nitrogenous material. Food proteid and tissue proteid, however, were the materials preferred by the organism, so ran the argument, and the large and incessant output of nitrogen which accompanied the intake of proteid was accepted as proof of the general truth of this idea. We might well question wherein lies the great advantage to the body in this continual excretion of nitrogen; whether the loss of energy in handling and removing the nitrogenous portion of the necessarily large proteid intake, in order to render available the non-nitrogenous part of the molecule, might not more than compensate for the supposed gain? But the truly astonishing fact that the output of nitrogen runs parallel with the intake of proteid, that the body cannot store up nitrogen to any large extent, has been taken as conclusive evidence that the organism prefers to use proteid for all of its requirements. Truly, we might just as well argue that this significant rise in the excretion of nitrogen after partaking of a proteid meal is an indication that the body has no need of this excess of nitrogen; that it is indeed a possible source of danger, since the system strives vigorously to rid itself of the surplus, and that the energy-needs of the body can be much more advantageously and economically met from fat and carbohydrate than from the carbonaceous residue resulting from the disruption of the proteid molecule.
In discussing these questions, we shall need to refer to several of the current theories concerning proteid metabolism, notably, the theories of Voit, Pflüger, and Folin. In 1867134 Carl Voit,50 of Munich, advanced the view that the proteid material of the body exists in two distinct forms, viz., as “morphotic” or “organized” proteid, representing proteid which has actually become a part of the living units of the body, i. e., an integral part of the living tissues; and “circulating” proteid, or that which exists in the internal meshes of the tissue, or in the surrounding lymph and circulating blood. The real point of distinction here is that while one portion of the body proteid is raised to the higher plane of living matter, i. e., becomes a component part of the living protoplasm, another and perhaps larger portion is outside of the morphological framework of the tissue, constituting a sort of internal medium which bathes the living cells, and acts as middleman between the blood and lymph on the one side and the living cells on the other. According to Voit’s view, it is this circulating proteid that undergoes metabolism; the proteid of the food after digestion and absorption being carried to the different tissues and organs, and then, without becoming an integral part of the living protoplasm of the cells, it is broken down under the influence of the latter. Obviously, small numbers of tissue cells are constantly dying, their proteid matter passing into solution, where it likewise undergoes metabolism. In other words, according to Voit, the great bulk of the proteid undergoing katabolism is the circulating proteid, derived more or less directly from the food, and which at no time has been a part of the tissue framework; while a smaller, but more constant amount, represents the breaking down of tissue cells. This conception of proteid metabolism is akin to our conception of morphological and physiological destruction. In the words of Foster: “We know that an epithelial cell, as notably in the case of the skin, may be bodily cast off and its place filled135 by a new cell; and probably a similar disappearance of and renewal of histological units takes place in all the tissues of the body to a variable extent. But in the adult body these histological transformations are, in the cases of most of the tissues, slow and infrequent. A muscle, for instance, may suffer very considerable wasting and recover from that wasting without any loss or renewal of its elementary fibres. And it is obvious that the metabolism of which we are now speaking does not involve any such shifting of histological units. On the other hand, we find these histological units, the muscle fibre or the gland cell, for instance, living on their internal medium, the blood, or rather on the lymph, which is the middleman between themselves and the actual blood flowing in the vascular channels.”
Voit claims that the proteid dissolved in the fluids of the body is more easily decomposable than that which exists combined in organized form, or as more or less insoluble tissue proteid; and it is this soluble and circulating form which, under the influence of the living cells, undergoes destruction or metabolism. We know, as has been previously stated, that oxidation does not take place to any extent in the circulating blood, and similarly there is every reason for believing that proteid metabolism does not occur in this menstrum. Metabolism is limited mainly to the active tissues of the body, but according to the present conception of the matter it does not occur at the expense of the proteid of the living cells, but involves material contained in the fluids bathing the cells; i. e., it is not the organized proteid that undergoes metabolism, but the proteid circulating in and about the internal meshes of the cells and tissues, the living cell being the active agent in controlling the process. Further, this view lessens the difficulty of understanding the elimination of nitrogen after a meal rich in proteid. If it was necessary to assume that all the proteid136 of our daily food is built up into living protoplasm before katabolism occurs, it would be exceedingly difficult to explain the sudden and rapid elimination of nitrogen which follows the ingestion of proteid. For example, we can hardly imagine that merely eating an excess of proteid food will lead to an actual breaking down of the living framework of the tissues, equivalent to the amount of nitrogen which the body at once eliminates. Voit’s theory, on the other hand, supposes a twofold origin of the nitrogen excreted; one part, the larger and variable portion, comes from the direct metabolism of the circulating proteid, being the immediate result of the ingested food and varying in amount with the quantity of proteid food consumed; the other, smaller and less variable in amount, has its origin in the metabolism of the true tissue proteid, or the actual living framework of the body.
In a fasting animal, the tissues and organs of the body still contain a large proportion of proteid matter, yet only a small fraction of this proteid is eliminated each day, hardly 1 per cent. If, however, proteid is absorbed from the intestine, proteid metabolism is at once increased, and the excretion of nitrogen may be fifteen times greater than during hunger. In other words, the extent of proteid metabolism is not at all proportional to the total amount of proteid contained in the body as a whole, but runs parallel in a general way with the quantity of proteid absorbed from the intestine. Obviously, the newly absorbed proteid is quite different in nature from the proteid which in much larger amounts is deposited throughout the body, since it is not organized and is so much more easily decomposable (Voit). This is the circulating proteid of the body; it exists in solution, and it is a significant fact that, according to Voit, the chemical transformations that characterize proteid katabolism occur only in solution. The organized proteid, on the other hand, is in137 a state of suspension, and its katabolism, which is relatively very small, is preceded by solution of the proteid in the fluids of the tissue, after which its further breaking down is assumed to be the same as that of the circulating proteid. This latter view is a fundamental part of the Voit theory; in long-continued fasting, for example, the living protoplasm of the various tissues and organs is of necessity drawn upon for the nourishment of the more vital parts of the body, such as the brain, spinal cord, etc., consequently the organized proteid is gradually dissolved and then decomposed, after it has become liquefied and has thus lost its organized structure.
In this conception of proteid metabolism, we picture the different organs and tissues of the body as being permeated by a fluid which carries variable amounts of nutritive material, the quantity of the latter determining in a way the extent of the proteid katabolism which shall take place. As the proteid of the food passes into the blood and lymph, the fluids bathing the cells are correspondingly enriched, and as a result, proteid katabolism is accelerated in parallel degree. During hunger, on the other hand, the organized proteid of the tissue cells is gradually liquefied and passes out into the current of the circulating fluids. As before stated, the organized proteid as such is never decomposed; it must first enter into solution, and then under the influence of the living cells it undergoes disruption in the same manner as the circulating proteid. It is thus evident that the tissue cells and the circulating fluids permeating them bear an ever changing relationship to each other. Excess of circulating proteid will be attended by increased katabolism, while at the same time there may be some accumulation of proteid in the cells, and indeed some conversion into organized proteid. During fasting, hunger, or with an insufficient intake of proteid food, the current will naturally be in the opposite direction, and organized proteid will slowly, but surely, be drawn upon.
138
Again, we may ask in view of these facts, of what real use to the body is this large katabolism of circulating proteid? We can easily understand the need of proteid to supply the loss incidental to the breaking down of organized or true tissue proteid, but this we are led to believe is very small in amount. Is there any real need for proteid beyond this requirement? The physiological fuel value of proteid is no greater than that of carbohydrate and considerably less than half that of fat, consequently there is on the surface no apparent reason why proteid should be used for its energy value in preference to the non-nitrogenous foodstuffs. Further, as we have seen, the energy of muscle work comes mainly, at least, from the breaking down of fat and carbohydrate; proteid, in the case of the well-nourished individual, ordinarily playing no part in this important line of energy exchange. Lastly, in the katabolism of proteid there is the large proportion of nitrogenous matter to be split off and disposed of before the carbon moiety of the molecule can be rendered available. Here, we have involved not only a loss of energy, but in addition a certain amount of what appears to be useless labor thrown upon the liver, kidneys, and other organs. Is there any wonder that the thoughtful physiologist, looking at the facts and theories presented by the Voit conception of proteid katabolism, should ask wherein lies the value to the body of this high rate of metabolism of circulating proteid, a rate of metabolism which is seemingly governed primarily by the amount of proteid food ingested?
Turning next to Pflüger’s51 views regarding proteid katabolism, we find a totally different outlook. Here, the supposition prevails that the plasma of the blood and lymph, with its contained proteid, is the food of the organs or their cells,139 but that before this food material can undergo katabolism it must first be absorbed by the cell and built up into the living protoplasm of the tissue. In other words, according to the views expressed by Pflüger, katabolism must be preceded by organization of the proteid. Expressed in still different language, the proteid material circulating in blood and lymph must be eaten up by the hungry cells and, by appropriate anabolic processes, made an integral part of the living protoplasm before disassimilation can occur. Further, according to Pflüger’s conception of these processes, there is a radical difference in the chemical nature of living protoplasm as compared with that of the so-called circulating proteid. The latter is looked upon as being comparatively stable, resisting oxidation in high degree, and hence not prone to undergo metabolism. Living protoplasm, on the other hand, is characterized by instability, suffering oxidation with the greatest ease, and hence readily broken down in the ordinary processes of katabolism. Assuming for the moment the correctness of this theory, we see at a glance that all disruption of proteid matter in the body must be preceded by the upbuilding of the proteid into living protoplasm. There can be no destruction of proteid until the latter has been raised to the high plane of living matter. The dead, inert circulating proteid can serve simply as food for the living cells, and cannot undergo katabolism until it has been built up into the organized structure of the tissue or organ. Even though we grant that a small proportion of proteid may suffer katabolism without previous organization, it does not materially modify the general trend of the argument that, according to Pflüger’s hypothesis, proteid katabolism is essentially a process involving the disruption of living protoplasm.
Consider what this means in the light of facts already presented. Remembering that the one factor above all others influencing the rate of proteid katabolism is the amount of140 proteid food taken in, and that the output of nitrogen, no matter what the previous condition of the body or the amount of proteid food ingested, runs more or less parallel with the consumption of proteid, we are forced to the conclusion, in accepting this hypothesis, that there must be superhuman activity in the building up of living protoplasm, only to be followed, however, by its immediate and more or less complete breaking down. Further, think of the daily or periodical fluctuation in the construction of bioplasm, coincident with variations in the amount of proteid food consumed, and the corresponding destruction of bioplasm as indicated by the daily output of nitrogen. Imagine, if you will, the concrete case of a man of 70 kilos body-weight eating a daily ration containing 125 grams of proteid, the nitrogen equivalent of which is practically excreted within twenty-four hours, and are we not wise in hesitating to believe that all of that proteid has been so quickly built up into living or organized tissue only to be immediately broken down and thrown out of the body? Think of the enormous activity implied in the manufacture of this bioplasm in the time allotted, and for what? Apparently, so that it can be broken down again. But such energy as is liberated in the breaking-down process might be derived far more economically by simple destruction of the proteid, as contained in the meshes of the tissue elements, without assuming a preliminary conversion into living protoplasm. Obviously, we have here a theory which does not help us in arriving at any very satisfactory conception of proteid metabolism. The facts which Pflüger and his followers bring forward in support of the theory are not very convincing, or at least not sufficiently so to carry conviction in the face of a natural disinclination to believe in the necessity of such a profound anabolic process, merely as a prelude to the speedy destruction of the finished product. Finally, we may add141 that if all proteid katabolized in the body must first be raised to the high level of living protoplasm before the final disruption can occur, it may be prudent to keep the daily intake of this foodstuff down to a level somewhat commensurate with the real needs of the body.
As has been stated many times in the course of this presentation, the most striking feature of proteid metabolism is the rapidity with which large quantities of proteid consumed as food are broken down, and the contained nitrogen eliminated from the body as urea. A few hours will suffice to accomplish the more or less complete destruction of food proteid; and any theory of proteid metabolism, to be at all satisfactory, must explain this peculiar phenomenon. According to recent investigations, it seems probable that some, at least, of the cleavage products of proteid formed during intestinal digestion are not built up into new proteid, but are at once eliminated mainly in the form of urea, without becoming a part of either the so-called circulating proteid, or the living protoplasm of the body. It will be recalled that under the influence of the digestive enzymes, trypsin and erepsin, proteid foodstuffs may be broken down while undergoing intestinal digestion into monamino- and diamino-acids, such as leucin, tyrosin, arginin, lysin, etc. A certain proportion of these comparatively simple substances may be directly absorbed by the portal circulation and carried to the liver, where they may undergo conversion into urea. In this way, some portion of the nitrogen of the ingested food may be quickly eliminated from the system. As has been stated in another connection, we are not sure at present how far proteid decomposition of the kind indicated takes place normally in the body. We merely know that there are present in the intestine, enzymes capable of splitting up proteid into these small fragments, and that substances of this type when made to circulate through the liver are transformed into urea. These142 facts, coupled with the well-known tendency of the nitrogen of proteid food to appear in the excretions a few hours after the food in question has been consumed, naturally suggests a direct breaking down of proteid along the lines indicated, with a possible retention of a carbonaceous residue (nitrogen-free) for subsequent oxidation, as a source of energy for heat or work. Obviously, all of the proteid food cannot behave in this manner, for if such were the case there would be no proteid available for making good the normal waste incidental to tissue changes. Either a certain amount of proteid escapes this profound alteration produced by the proteolytic enzymes in question, or else a certain proportion of these simple decomposition products is synthesized in the intestine, or in the tissues of the body, to form new proteid for the regeneration of cell protoplasm. However this may be, we have presented in this view a plausible explanation of the prompt appearance of food nitrogen in the excretions, and without compelling belief in a theory, such as Pflüger’s, which taxes one’s credulity to the utmost. To be sure, as a prominent writer on physiology has recently said, such a view stands opposed to our conceptions of the importance of proteid food; but it seems possible, in the light of accumulating knowledge, that our conceptions of the part played by proteid foods in the nutrition of man have not been strictly logical, or quite in accord with true physiological reasoning.
Again, in this connection, we may ask the question, why is it that the body provides such an effective method for the speedy breaking down of proteid food and the prompt elimination of the contained nitrogen? Whatever the means made use of by the organism in accomplishing this, the result is the same; the nitrogen of the ingested food is, in large measure, quickly gotten rid of. We clearly recognize the all-important position of proteid foods in the nutrition of the body, but there appears a certain inconsistency in this prompt removal143 of the nitrogen-containing portion of the proteid molecule. The nitrogenous part of the proteid food is, physiologically considered, the all-important part. It is the only source of nitrogen available to the system, and yet apparently the larger proportion of this nitrogenous material is not utilized in any recognizable way, but is eliminated as quickly as possible. Is it not within the limits of possibility that these methods, whatever may be the exact mechanism involved, are merely a means of getting rid of a surplus of proteid for which the body has no real need? This question I shall try to answer later on in another connection, but we may advantageously keep this possibility in mind while we are discussing these theories of proteid metabolism.
It is obvious, in the light of present knowledge, that there must be a certain amount of true tissue proteid broken down each day, independent of that larger metabolism coincident with the intake of proteid food. However much this more voluminous proteid katabolism may fluctuate, owing to variations in the intake of proteid, and whatever the significance of this latter phase of metabolism, it is self-evident that there must be a steady, constant metabolism, upon which the life of the various tissues and organs of the body depends, and by which the proteid integrity of the tissue cells is maintained. This implies a certain degree of true tissue change, in which definite amounts of proteid material are broken down and the resultant loss made good from the proteid intake. No matter what specific name be applied to this form of proteid katabolism, its existence is clearly recognized. It is obviously a form of metabolism distinct, and probably quite different, from that form, more variable in extent, which is associated with the intake of proteid food. Plainly, if there is truth in these statements, there should be some data available by means of which these two lines of proteid katabolism can be more or less sharply differentiated.
144
Thanks especially to the work of Folin,52 these data are now apparently at hand, and the facts which he has accumulated with painstaking care seem destined to throw additional light upon our conception of proteid metabolism. It will be remembered that in the breaking down of proteid, the great bulk of its contained nitrogen is eliminated in the form of urea. In addition, a certain smaller amount of nitrogen is excreted in the forms of creatinin and uric acid. As we have seen, the total output of nitrogen, which measures the extent to which proteid is decomposed in the body, varies with the intake of proteid food; but it is found that the proportion of nitrogen excreted in the forms of urea and uric acid varies with the extent of the metabolism. In other words, quantitative changes in the daily proteid katabolism are accompanied by pronounced changes in the distribution of the excreted nitrogen. Let us take a single illustration from Folin’s results; the case of a healthy man who on one day—July 13—consumed a proteid-rich diet, and on the other day—July 20—was living on a diet containing only about 1 gram of nitrogen. The composition of the excretion through the kidneys on these two days is shown in the following table:
July 13. |
July 20. |
|
|---|---|---|
| Volume of urine | 1170 c.c. | 385 c.c. |
| Total nitrogen | 16.80 grams | 3.60 grams |
| Urea-nitrogen | 14.70 " = 87.5% | 2.20 " = 61.7% |
| Uric acid-nitrogen | 0.18 " = 1.1% | 0.09 " = 2.5% |
| Creatinin-nitrogen | 0.58 " = 3.6% | 0.60 " =17.2% |
Here we see, as would be expected, that on the high proteid diet, there was a large excretion of total nitrogen and of urea; while on the low proteid diet, nitrogen and urea were correspondingly diminished. The point to attract our attention, however, is the marked difference in the percentage of urea-nitrogen in the two cases; a difference which amounts to about 26 per cent. A similar difference is to be noted in the percentage of uric acid-nitrogen. Lastly, it is to be observed that in spite of the great difference in the extent of metabolism on the two days—an excretion of 16.8 grams of nitrogen, as contrasted with 3.6 grams—the amount of creatinin-nitrogen is essentially the same. Folin finds that these peculiarities in the percentage distribution of excreted nitrogen hold good in all cases where there is this wide divergence in the amount of proteid katabolized, and, further, that there is a gradual and regular transition from the one extreme to the other. He sees in these results evidence that there are in the body two forms of proteid katabolism, essentially independent and quite different. One kind is extremely variable in quantity, while the other tends to remain constant. The variable form has its own particular kind of waste products, of which urea is the chief. The constant katabolism, on the other hand, is largely represented by creatinin and to a lesser degree by uric acid. The more the total katabolism is reduced, the more prominent become creatinin and uric acid, products of the constant katabolism; while urea, as chief representative of the variable katabolism, becomes less conspicuous. Folin suggests the term endogenous or tissue metabolism for the constant variety, while the variable form he would name exogenous or intermediate metabolism.
In these suggestions we have not theory only, but a number of very important facts which plainly must have some significance. Take, for example, the excretion of146 creatinin. It is a characteristic nitrogenous waste product, but its elimination from the body is wholly independent of quantitative changes in the total amount of nitrogen excreted. In other words, the amount of creatinin eliminated is a constant quantity for a given individual under ordinary conditions, no matter how great the variation in the amount of proteid food, provided no meat is eaten. Meat must be avoided in testing this point, since meat contains a certain amount of creatin, or other components, which would be excreted as creatinin. Further, it is found that every individual has his own specific creatinin excretion, which fact again emphasizes the idea that this substance is a product of true tissue katabolism, having no connection with that variable metabolism, of which urea is the striking representative. These are facts which cannot be ignored. They are well established by the careful observations of Folin, and they are confirmed by a large number of observations made in our own laboratory. Turn now to that other, more conspicuous, product of proteid katabolism, urea. With a so-called average proteid intake, about 88–90 per cent of the excreted nitrogen will be in the form of urea, but, as Folin states, “with every decided diminution in the quantity of total nitrogen eliminated, there is a pronounced reduction in the per cent of that nitrogen represented by urea. When the daily total nitrogen elimination has been reduced to 3 grams or 4 grams, about 60 per cent of it only is in the form of urea.” Here, we have the chief product of exogenous metabolism, a substance quite distinct from creatinin, just as the process by which it originates is likewise quite distinct.
Exogenous metabolism is plainly a process of quite a different order from that of endogenous, or tissue metabolism. The latter involves oxidation, while the former consists essentially of a series of hydrolytic cleavages which result in a rapid elimination of the proteid-nitrogen as urea. In this147 conception of exogenous katabolism, we have essentially the same viewpoint as was previously taken in attempting to explain how excess of proteid food can be so quickly decomposed, and its nitrogen removed from the body. Whether the hydrolytic cleavage is accomplished solely by trypsin and erepsin, whether it takes place only in the intestine and in the liver, or whether other glands and tissues are involved, is at present immaterial; the essential point is that we have in the body a variety of proteid katabolism, quite different from true tissue katabolism, the extent of which is dependent primarily upon the amount of proteid food consumed. The process involved is one which aims at the rapid removal of the proteid-nitrogen as urea; without incorporation of the absorbed proteid, or its decomposition products, either as an integral or adherent part of the tissue proteid. Hydrolytic cleavage is eminently fitted to accomplish this with the least expenditure of energy, while the carbonaceous residue of the proteid thus freed from nitrogen can be transformed into carbohydrate, or directly oxidized as the needs of the body demand.
As one considers these views so admirably worked out by Folin, the question naturally arises, if the real demands of the body for proteid food will not be adequately met by the quantity necessary to satisfy the true tissue metabolism? We may well believe, with Folin, that “only a small amount of proteid, namely, that necessary for the endogenous metabolism, is needed. The greater part of the proteid furnished with so-called standard diets, like Voit’s, i. e., that part representing the exogenous metabolism, is not needed; or, to be more specific, its nitrogen is not needed. The organism has developed special facilities for getting rid of such excess of nitrogen, so as to get the use of the carbonaceous part of the proteid containing it.” In endogenous metabolism, we have a steady, constant process quite independent of the amount of proteid food, and absolutely indispensable for the maintenance148 of life. So far as we know at present, its representative creatinin is, for a given individual, the same in amount during fasting as when a rich, meat-free, proteid diet is taken. The one factor that seemingly determines the amount of creatinin eliminated is the weight of the individual, or more exactly the weight of the true tissue elements of the body, as distinct from fat or adipose tissue. Endogenous or tissue katabolism obviously calls for a certain quantity of proteid to maintain equilibrium, but this is small in amount as compared with the usual intake of proteid foods. The average man, with his ordinary dietetic habits, consumes more nitrogen than the body can possibly make use of. The excess is not stored up, “because the actual need of nitrogen is so small that an excess is always furnished with the food, except, of course, in carefully planned experiments” (Folin).
We have seen at what low levels of proteid intake, nitrogen equilibrium can be established, and we may well have faith in the conception of an endogenous proteid katabolism which involves only minimal quantities of proteid. Further, we have observed the constant tendency of the body to maintain a condition of nitrogenous equilibrium, even with varying income, and how slow the body is to lay by nitrogen on a rich proteid diet, even when long deprived of proteid food; a fact difficult of explanation except on the assumption that the real need of the body for nitrogen is small, and that the tissues habitually carry a relatively large reserve of nitrogenous material. We may assume with Folin that “all the living protoplasm in the animal organism is suspended in a fluid very rich in proteid, and on account of the habitual use of more nitrogenous food than the tissues can use as proteid the organism is ordinarily in possession of approximately the maximum amount of reserved proteid in solution that it can advantageously retain. When the supply of food proteid is stopped, the excess of reserve proteid inside the organism is149 still sufficient to cause a rather large destruction of proteid during the first day or two of proteid starvation, and after that the proteid katabolism is very small, provided sufficient non-nitrogenous food is available. But even then, and for many days thereafter, the protoplasm of the tissues has still an abundant supply of dissolved proteid, and the normal activity of such tissues as the muscles is not at all impaired or diminished. When 30 grams or 40 grams of nitrogen have been lost by an average-sized man during a week or more of abstinence from nitrogenous food the living muscle tissues are still well supplied with all the proteid they can use. That this is so, is indicated on the one hand by the unchanged creatinin elimination, and on the other by the fact that one experiences no feeling of unusual fatigue or of inability to do one’s customary work. Because the organism at the end of such an experiment still has an abundance of available proteid in the nutritive fluids, it is at once seemingly wasteful with nitrogen when a return is made to nitrogenous food. This is why it only gradually, and only under the prolonged pressure of an excessive supply of food-proteid again acquires its original maximum store of this reserve material.”
We may reasonably suppose that the reserve of proteid present in the body is contained in the fluid media, and not as a part of the living protoplasm. Further, we are apparently justified in the belief that the sole form of proteid katabolism which is vitally important for the welfare of the body is the endogenous katabolism. This must be provided for adequately and indeed liberally, and in addition there should be sufficient intake to keep up an abundant supply of reserve proteid, but beyond these necessities there would seem to be no legitimate demand for additional proteid. The voluminous exogenous proteid katabolism so conspicuous in most individuals would seem to have no justification in fact,150 or in physiological reasoning. What good, for example, can be accomplished by this constant splitting off of nitrogen, with its subsequent speedy removal from the body? The organism can neither use it nor store it up, and why therefore should this daily burden of an excessive and accelerated proteid katabolism be borne? As we have seen, the energy of muscle work is derived mainly, and can come wholly, from the breaking down of non-nitrogenous materials, fats and carbohydrates. The very fact that an intake of say 120 grams of proteid is followed at once by the removal of the larger part of the contained nitrogen, as a result of the exogenous katabolism of the body, would seemingly warrant the view that the proteid so decomposed might advantageously be replaced by a corresponding amount of carbohydrate. In muscle work, as in heat production, carbohydrate and fat are the materials burned up, or oxidized. Proteid, on the other hand, is not so oxidized, at least not the nitrogen-containing portion of the molecule.
There are apparent only two possible reasons for assuming a need on the part of the body for the high exogenous katabolism of proteid so commonly observed. The one is that the carbonaceous residue left after the cleavage of nitrogen from the proteid molecule is better adapted for the needs of the body than either carbohydrate or fat. Although this does not seem very probable, it is of course a possibility and merits consideration. Feeding experiments, with a comparatively small proteid intake, continued over a sufficient length of time, would show conclusively how much weight should be attached to this hypothesis. The other possibility is that the body may derive some advantage from the presence, in the tissues and fluids, of the varied nitrogenous cleavage products split off from proteid so abundantly in exogenous katabolism. These substances are mainly amino-acids on their way to urea, and there is no apparent reason151 why they should be of service to the organism. Still, the processes going on in the tissues and organs of the body are intricate and not wholly understood, and we can conceive of some useful function of which as yet we have no knowledge. In the construction of tissue proteid, for example, as in a possible synthesis out of the fragments formed by hydrolytic cleavage, it is not impossible that certain corner-stones are needed, and that in order to obtain these there must be a more or less wasteful breaking down of food-proteid. However improbable this may seem, it, like the preceding hypothesis, can be tested in a way by adequate feeding experiments, which shall determine the effect on the body of a low proteid intake continued over a long period of time. On the other hand, it is equally plausible, and for some reasons more probable, to assume that this excessive exogenous katabolism may be in a measure prejudicial to the best interests of the body; that the many nitrogenous fragments formed in the efforts of the organism to prevent undue accumulation of reserve proteid may in the long run do as much harm as good.
Further, there is reason in the question whether the continual carrying of excessive amounts of nitrogen reserves in the shape of soluble proteid in the blood and lymph, and in the meshes of tissue and cell protoplasm, is advantageous for the maintenance of the highest degree of efficiency? We all recognize that an excessive accumulation of fat is distinctly disadvantageous to the welfare of the body, and there is, physiologically speaking, equally good ground for considering that the storage of unorganized proteid in amounts beyond all possible requirements of the body may be equally undesirable. Because less tangible to the eye, the accumulation of unnecessary proteid is not so easily recognizable, but this fact does not diminish the possible danger which such accumulation may constitute. It must be granted, however,152 that we are dealing here with hypotheses and not facts, but though hypothetical the suggestions made are of sufficient moment to merit attention and experimental study. In a later chapter, we shall have occasion to present some facts bearing on these questions.
In the meantime, we may lay due stress upon the significance of these views regarding proteid katabolism. We must accept as settled the general idea that there are two distinct forms of proteid katabolism within the body; one form representing the decay of tissue or cell protoplasm, small in amount, with its own particular decomposition products, and absolutely essential for the continuance of life. The other form, the so-called exogenous katabolism, runs a totally different course with distinctive side-products and end-products; it is variable in extent, in harmony with variations in proteid intake, and subject to the suspicion that at the level ordinarily maintained it constitutes a menace to the preservation of that high degree of efficiency which is an attribute of good health.
153
DIETARY HABITS AND TRUE FOOD REQUIREMENTS
Topics: Dietetic customs of mankind. Origin of dietary standards. True food requirements. Arguments based on custom and habit. Relationship between food consumption and prosperity. Erroneous ideas regarding nutrition. Commercial success and national wealth not the result of liberal dietary habits. Instinct and craving not wise guides to follow in choice and quantity of food. Physiological requirements and dietary standards not to be based on habits and cravings. Old-time views regarding temperate use of food. The sayings of Thomas Cogan. The teachings of Cornaro. Experimental results obtained by various physiologists. Work of the writer on true proteid requirements. Studies with professional men. Nitrogen equilibrium with small amounts of food. Sample dietaries. Simplicity in diet. Nitrogen requirement per kilogram of body-weight. Fuel value of the daily food. Experiments with university athletes. Nitrogen balance and food consumption. Sample dietaries. Adequacy of a simple diet.
Having acquired information regarding the principles of metabolism and the general laws governing the nutrition of the body, we may next consider briefly the dietetic habits of mankind, with a view to learning how far such habits coincide with actual nutritive requirements. Eventually, we shall need to ask the questions: What are the true nutritive requirements of the body? How much food and what kinds of food does the ordinary individual doing an average amount of work need each day in order to preserve body equilibrium, and to maintain health, strength, and vigor under the varying conditions of life? What amount of nitrogen or proteid, and what the total calorific value154 required to supply the physiological needs of the body? How closely do the so-called “normal diets” and “standard diets,” which have met with such general acceptance, conform to a rational conception of true physiological needs? These are vital questions of great physiological and economic importance, and they are not easily answered; but theoretical considerations based on scientific data, and experimental evidence combined with practical experience, should point the way to some very definite conclusions.
Observations made in many countries regarding the dietetic customs and habits of the people have resulted in the establishment of certain dietary standards, which have been more or less generally adopted as representing the requirements of the body. As a prelude to the discussion of this question, let us consider briefly some of the results of these dietary studies. In Sweden, laborers doing hard work were found by Hultgren and Landergren to consume daily, on an average, 189 grams of proteid, 714 grams of carbohydrate, and 110 grams of fat, with a total fuel value for the day’s ration of 4726 large calories. In Russia, workmen at moderately hard labor, having perfect freedom of choice in their food, were found by Erisman to take daily 132 grams of proteid, 584 grams of carbohydrate, and 79 grams of fat, this ration having a fuel value of 3675 calories. In Germany, soldiers in active service consumed daily, according to Voit, 145 grams of proteid, 500 grams of carbohydrate, and 100 grams of fat, with a fuel value of 3574 calories. In Italy, laborers doing a moderate amount of work were found by Lichtenfelt to consume daily 115 grams of proteid, 696 grams of carbohydrate, and 26 grams of fat, with a fuel value of 3655 calories. In France, Gautier states that the ordinary laborer working eight hours a day must have 135 grams of proteid, 700 grams of carbohydrate, and 90 grams of fat daily, with a fuel value of 4260 calories. In England, weavers were found to take155 daily 151 grams of proteid, with carbohydrates and fats sufficient to make the total fuel value of the day’s ration equal to 3475 calories. In Austria, farm laborers consumed daily 159 grams of proteid, with carbohydrates and fats sufficient to raise the fuel value of the food to 5096 calories.
Subjects. |
Proteid consumed Daily. |
Total Fuel Value of Daily Food. |
|---|---|---|
grams |
calories |
|
| Swedish laborers, at hard work | 189 |
4726 |
| Russian workmen, moderate work | 132 |
3675 |
| German soldiers, active service | 145 |
3574 |
| Italian laborers, moderate work | 115 |
3655 |
| French laborers, eight hours’ work | 135 |
4260 |
| English weavers | 151 |
3475 |
| Austrian farm laborers | 159 |
5096 |
| American Subjects. | ||
| Man with very hard muscular work | 175 |
5500 |
| Man with hard muscular work | 150 |
4150 |
| Man with moderately active muscular work |
125 |
3400 |
| Man with light to moderate muscular work |
112 |
3050 |
| Man at “sedentary” or woman with moderately active work |
100 |
2700 |
Observations of this order might be multiplied indefinitely, but the above will suffice to give a general idea of the average food consumption of European peoples doing a moderate amount of work. These data, however, must be supplemented by the observations made in our own country, which have been very extensive, through the “investigations on the nutrition of man in the United States,” carried on by the Office of Experiment Stations in the Department of Agriculture, under the efficient leadership of Atwater. As stated by Messrs.156 Langworthy and Milner, in an official bulletin issued in 1904, dietary studies of the actual food consumption of people of different classes in different parts of the United States have been made during the years 1894 to 1904 on about 15,000 persons,—men, women, and children,—as a result of which it is indicated that “the actual food requirements of persons under different conditions of life and work” vary from 100 to 175 grams of proteid per day, with a total fuel value ranging from 2700 to 5500 calories. For comparison, the various data may be tabulated as shown on page 155.
These figures by no means represent maximum food consumption. Thus, studies have been made on fifty Maine lumbermen,53 where the intake of proteid food averaged 185 grams per day, with a total fuel value of 6400 calories. Further, dietary studies of university boat crews54 have shown fairly high results. The Yale University crew, while at Gales Ferry, averaged per man during seven days 171 grams of proteid, 171 grams of fat, and 434 grams of carbohydrate, with a total fuel value of 4070 calories per day. The members of the Harvard University crew showed an average daily consumption of 160 grams of proteid, 170 grams of fat, and 448 grams of carbohydrate, with a total fuel value of 4074 calories. It is also reported that a football team of college students in the University of California consumed daily, per man, 270 grams of proteid, 416 grams of fat, and 710 grams of carbohydrate, with a total fuel value of 7885 calories. These figures may be contrasted, however, with the data obtained in a study of the dietary habits of fourteen professional men’s families, where the average amount of proteid consumed daily was 104 grams, fat 125 grams, and carbohydrate 423 grams, with a total fuel value of 3325 calories.
157
Leaving out of consideration the extremes given, it is undoubtedly true that, within certain rather wide limits, there is an apparent tendency for people of different nations, having a free choice of food and not restricted by expense, to consume daily approximately the same amounts of nutrients; to use what may be called liberal rather than small amounts of food; and, lastly, to consume food somewhat in proportion to the amount of work done. It is perhaps, therefore, not strange that students of nutrition should have taken these results, obtained by the statistical method, as indicating the actual needs of the body for food, and that so-called “standard diets” and “normal diets” should have been constructed, based upon these and corresponding data. Thus, we have the widely adopted “Voit standard,” composed of proteid 118 grams, carbohydrate 500 grams, and fat 56 grams, with a total fuel value of 3055 calories, as the amount of food required daily by a man of 70 kilos body-weight doing a moderate amount of work. These figures were obtained by Voit as an average of the food consumption of a large number of laboring men in Germany, and they carried additional weight because at that time Voit and others thought they had evidence that nitrogenous equilibrium could not be maintained for any length of time on smaller amounts of proteid.
The figures given in the preceding table under the head of American subjects constitute the “Atwater standards,” and as already indicated, are based upon the dietetic habits of over 15,000 persons under different conditions of life and physical activity. In the words of the official Bulletin, these standards covering the quantities of food per day “are intended to show the actual food requirements of persons under different conditions of life and work.” Here, however, lies an assumption which seems to meet with wide acceptance, but for which it is difficult to conceive any158 logical reason. The thousands of dietary studies made on peoples all over the world, affording more or less accurate information regarding the average amounts of proteid, fat, and carbohydrate consumed under varying conditions, are indeed most interesting and important, as affording information regarding dietetic customs and habits; but, the writer fails to see any reason why such data need be assumed to throw any light on the actual food requirements of the body. In the words of another, “Food should be ingested in just the proper amount to repair the waste of the body; to furnish it with the energy it needs for work and warmth; to maintain it in vigor; and, in the case of immature animals, to provide the proper excess for normal growth, in order to be of the most advantage to the body” (Benedict).
Any habitual excess of food, over and above what is really needed to meet the actual wants of the body, is not only uneconomical, but may be distinctly disadvantageous. Voit, among others, has clearly emphasized the general principle that the smallest amount of proteid, with non-nitrogenous food added, that will suffice to keep the body in a state of continual vigor is the ideal diet. My own conception of the true food requirements of the body has been expressed in the statement that man needs of proteids, fats, and carbohydrates sufficient to establish and maintain physiological and nitrogen equilibrium; sufficient to keep up that strength of body and mind that is essential to good health, to maintain the highest degree of physical and mental activity with the smallest amount of friction and the least expenditure of energy, and to preserve and heighten, if possible, the ordinary resistance of the body to disease germs. The smallest amount of food that will accomplish these ends is, I think, the ideal diet. There must truly be enough to supply the real needs of the body, but any great surplus over and above what is actually called for may in the long run prove an undesirable addition.159 With these thoughts in mind, may we not reasonably ask why it should be assumed that there is any tangible connection between the dietetic habits of a people and their true physiological needs?
Arguments predicated on custom, habit, and usage have no physiological basis that appeals strongly to the impartial observer. Man is a creature of habits; he is quick to acquire new ones when his environment affords the opportunity, and he is prone to cling to old ones when they minister to his sense of taste. The argument that because the people of a country, constituting a given class, eat a certain amount of proteid food daily, the quantity so consumed must be an indication of the amount needed to meet the requirements of the body, is as faulty as the argument that because people of a given community are in the habit of consuming a certain amount of wine each day at dinner their bodies must necessarily be in need of the stimulant, and that consequently alcohol is a true physiological requirement. A large proportion of mankind is addicted to the tobacco habit, and to many persons the after-dinner cigar is as essential to comfort as the dinner itself; but would any one think of arguing that tobacco is one of the physiological needs of the body?
It is said that dietary studies made all over the civilized world “show that a moderately liberal quantity of protein is demanded by communities occupying leading positions in the world. . . . It certainly seems more than a remarkable coincidence that peoples varying so widely in regard to nationality, climatic and geographical conditions, and dietetic habits, should show such agreement in respect to consumption of protein and energy.” Again, we hear it said that “whatever may be true of a few individuals, with communities a generally low condition of mental and physical efficiency, thrift, and commercial success, is coincident with a low proportion of protein in the diet.” The writer, however, fails160 to find evidence in the results afforded by dietary studies that there is any causal relationship between the amount of proteid food consumed and the mental or physical supremacy of the people of a given nation or community. Cause and effect are liable to become reversed in arguments of this kind. It is certainly just as plausible to assume that increase in the consumption of proteid follows in the footsteps of commercial and other forms of prosperity, as to argue that prosperity or mental and physical development are the result of an increased intake of proteid food.
Proteid foods are usually costly, and the ability of a community to indulge freely in this form of dietetic luxury depends in large measure upon its commercial prosperity. The palate is an extremely sensitive organ, and the average individual properly derives great satisfaction from the pleasurable effects of tasty articles of food. Furthermore, there are many curious and quite unphysiological notions abroad regarding foods, which tend to incite persons to unnecessary excess and extravagance whenever they acquire the means to do so. The latter point is well illustrated by the more or less prevalent opinion that a cut of tenderloin steak is more nutritious than a cut of round steak. It is true that the former is apt to be more tender, to have a little finer flavor; but the round steak, when properly prepared, is just as nutritious, and equally capable of meeting the needs of the body, as the more expensive tenderloin. With increasing prosperity, we turn at once, as a rule, to the more tasty and appetizing viands, partly to satisfy the craving of appetite and palate, and partly because there is an inherent belief that these varied delicacies, accessible to the prosperous community, count as an aid to health and strength. The poor laborer, with his small wage, is restricted to a certain low level of dietary variety, and must likewise be economical as to quantity, but on the first opportunity afforded by a fuller161 purse he is apt to pass from corned beef to a fresh roast with its more appetizing flavor; to eschew brown bread in favor of the white loaf, and in many other ways to evince his desire for a dietary which, though perhaps no more nutritious, appeals because of its finer flavor, more appetizing appearance, and greater variety. He is in the same position as the smoker who, limited by his purse to a five-cent cigar after dinner, quickly passes to a cigar of better flavor as soon as his finances warrant the indulgence. At the same time, if prosperity continues, our laborer will speedily pass to a higher level of proteid intake and greater fuel value, through increased consumption of meat and butter, together with other articles rich in proteid and fat.
In this connection, we may emphasize a fact of some significance in its bearing on dietetic customs; viz., that ever since Liebig advanced his theory that proteid material is the sole source of muscular energy, there has been a deep-rooted belief that meat is the most efficient kind of food for keeping up the strength of the body, and hence especially demanded by all whose work is mainly physical. Although this view, as we have seen, has been thoroughly disproved, the idea is still more or less generally held that an abundance of meat is a necessary requisite for a good day’s work, a view which undoubtedly accounts in some measure for the tendency toward a high proteid intake, evinced by many of the laboring class whose means will permit the necessary outlay.
Increased consumption of proteid food may be coincident with thrift and commercial success, but there is no justification for the belief that these are the result of changed dietary conditions. The dietary of our New England forefathers was, according to all accounts, exceedingly limited as compared with that of to-day, but it is doubtful if the present generation is any better developed, physically or mentally,162 than the stalwart and vigorous people who opened up this country to civilization. To-day, as a nation, we have greater wealth, and our commercial prosperity has become phenomenal; but would any one think for a moment that these characteristics are attributable to the large consumption of proteid food so common to this generation of the American people? No, increased wealth simply paves the way for greater freedom in the choice of food; increased commercial success and business prosperity throw open avenues which formerly were closed; greater variety of animal foods, and vegetable foods as well, rich in proteid, are made easily accessible, and appeal to eye and palate on all sides; appetite and craving for food are abnormally stimulated, and dietetic habits and customs change accordingly. In the words of another, “the one thing that primitive, barbarous, and civilized man alike long for is an abundance of the ‘flesh-pots of Egypt.’ The very first use the latter makes of his increased power and financial resources is to buy new, rare, and expensive kinds of meat.” With these facts before us, it is difficult to accept the assumption that dietetic customs afford any indication of the food requirements of the body. To the physiologist such a view does not appeal, since there is a lack of any scientific evidence that carries conviction.
But it may be asked, is not appetite a safe guide to follow? Do not the cravings of the stomach and the so-called pangs of hunger merit consideration? Is it not the part of wisdom to follow inclination in the choice and quantity of our food? Can we not safely rely upon these factors as an index of the real needs of the body? If these questions are to be answered in the affirmative, then it is plain that a study of dietetic customs will tell us definitely how much food and what kinds of food are required daily to supply the true wants of the body. There are writers who claim that instinct is a perfectly safe guide to follow; that it is far superior to163 reason; but it is to be noticed that most of these writers, if they have any physiological knowledge to draw upon, are sooner or later prone to admit that the body has certain definite needs which it is the purpose of food to supply, with the added implication that any surplus of food over and above what is necessary to meet these demands is entirely uncalled for. Thus, one such writer states that “the man in the street follows his God-given instincts and plods peacefully along to his three square meals a day, consisting of anything he can find in the market, and just as much of it as he can afford, with special preference for rich meats, fats, and sugars.” Yet this same writer, a little later, emphasizes the fact that “every particle of the energy which sparkles in our eyes, which moves our muscles, which warms our imaginations, is sunlight cunningly woven into our food by the living cell, whether vegetable or animal. Every movement, every word, every thought, every aspiration represents the expenditure of precisely so much energy derived from food.” Why, then, would it not be wise to ascertain how much energy is so expended, on an average, during the day’s activity and govern the intake of food accordingly? Why not apply an intelligent supervision in place of following an instinct which, in the words of the author just quoted, leads one on to consume “anything he can find in the market and just as much of it as he can afford”? Truly, if dietetic customs and the habits of mankind are the results of instinct working in this fashion, there cannot be much value in the data obtained by observing the quantities of food mankind is in the habit of eating. Dietary standards based on such observations must be open to the suspicion of representing values far above the actual needs of the body.
Habits and cravings are certainly very unreliable indices of true physiological requirements. Man is constantly acquiring new habits, and these in time become second nature,164 forcing him to practise that which he has become accustomed to, regardless of whether it is beneficial or otherwise. The celebrated philosopher, John Locke, in his essay on education, says: “I do not think all people’s appetites are alike . . . but this I think, that many are made gourmands and gluttons by custom, that were not so by nature; and I see in some countries, men as lusty and strong, that eat but two meals a day, as others that have set their stomachs by a constant usage, like Larums, to call on them for four or five.” Again, the so-called cravings of appetite are largely artificial and mainly the result of habit. A habit once acquired and persistently followed soon has us in its grasp, and then any deviation therefrom is very apt to disturb our physiological equilibrium. The system makes complaint, and we experience a craving, it may be, for that to which the body has become accustomed. There has thus come about a sentiment that the cravings of the appetite for food are to be fully satisfied, that this is merely obedience to nature’s laws. In reality, there is no foundation for such a belief; any one with a little persistence can change his or her habits of life, change the whole order of cravings, thereby indicating that the latter are essentially artificial, and that they have no necessary connection with the welfare or needs of the body. The man who for some reason deems it advisable to adopt two meals a day in place of three or four, at first experiences a certain amount of discomfort, but eventually the new habit becomes a part of the daily routine, and the man’s life moves forward as before, with perfect comfort and without a suggestion of craving, or a pang of hunger. Dietetic requirements, and standard dietaries, are not to be founded upon the so-called cravings of appetite and the instinctive demands for food, but upon reason and intelligence, reinforced by definite knowledge of the real necessities of the bodily machinery.
165
The standards which have been adopted more or less generally throughout the civilized world, based primarily on the assumption that man instinctively and independently selects a diet that is best adapted to his individual needs, are open to grave suspicion. The view that the average food consumption of large numbers of individuals and communities must represent the true nutritive requirements of the people is equally untenable. Naturally, there is general recognition of the principle that food requirements are necessarily modified by a variety of circumstances, such as age, sex, body-weight, bodily activity, etc. It is obvious that the man of 140 pounds body-weight needs less proteid than the man of 170 pounds, and that the man who does a large amount of physical work demands a larger calorific value in his daily diet, i. e., more carbohydrate and fat, than the sedentary individual. The growing child, in proportion to his body-weight, plainly needs more proteid for the upbuilding of tissue, and there are many conditions of disease where special dietetic treatment is undoubtedly called for. Our contention, however, and one which we believe to be perfectly justifiable, is that the true food requirements of the body, under any conditions, cannot be ascertained with any degree of accuracy by observations of what people are in the habit of eating; that customs and habits are not a safe index of true physiological needs. On the contrary, we are inclined to the belief that direct physiological experimentation, covering a sufficient length of time and with an adequate number of individuals, will prove far more efficient in affording a true estimate of the quality and quantity of food best adapted for the maintenance of good health, strength, and vigor.
Before considering these latter points, it is interesting to note, in passing, that during the last four centuries many thoughtful men have called attention to the apparent excessive use of food. There seems to have been in many quarters166 a more or less prevalent opinion that custom and habit were leading people on to methods of living, which were not in accord with the best interests of the community. It must be remembered, however, that arguments of this kind, even fifty years ago, could have been founded only on general observation and the application of common sense, since there were then no sound physiological data on which to predicate an opinion, or base a conclusion. Still, there were men of authority who attempted to lay before the people a proper conception of the temperate use of food. We have not the time here to consider many of these pleas, but I venture to call attention to the somewhat celebrated book published by the physician Thomas Cogan in 1596, under the title “The Haven of Health,” and dedicated “to the right honorable and my verie good lord, Sir Edward Seymour, Knight and Earl of Hertford.” Under the subject of diet, this old-time writer says: “The second thing that is to be considered of meates is the quantitie, which ought of all men greatly to be regarded, for therein lyeth no small occasion of health or sickness, of life or death. For as want of meate consumeth the very substance of our flesh, so doth excesse and surfet extinguish and suffocate naturall heat wherein life consisteth.” Again, “Use a measure in eating, that thou maist live long: and if thou wilst be in health, then hold thine hands. But the greatest occasion why men passe the measure in eating, is varitie of meats at one meale. Which fault is most common among us in England farre above all other nations. For such is our custome by reason of plentie (as I think) that they which be of abilitie, are served with sundry sortes of meate at one meale. Yea the more we would welcome our friends the more dishes we prepare. And when we are well satisfied with one dish or two, then come other more delicate and procureth us by that meanes, to eate more than nature doth require. Thus varietie bringeth us to excesse, and sometimes167 to surfet also. But Phisicke teacheth us to faede moderately upon one kinde of meate only at one meale, or at leastwise not upon many of contrarie natures. . . . This disease, (I mean surfet) is verie common: for common is that saying and most true: That more die by surfet than by the sword. And as Georgius Pictorius saith, all surfet is ill, but of bread worst of all. And if nature be so strong in many, and they be not sicke upon a full gorge, yet they are drowsie and heavie, and more desirous to loyter than to labor, according to that old maeter, when the belly is full, the bones would be at rest. Yea the minde and wit is so oppressed and overwhelmed with excesse that it lyeth as it were drowned for a time, and unable to use his force.”
Cogan likewise makes some interesting statements regarding the effects of custom on the consumption of proteid food, especially meats. Quoting further from this author: “The fourth thing that is to be considered in meats is custome. Which is of such force in man’s bodie both in sicknesse and in health, that it countervaileth nature itselfe, and is therefore called of Galen in sundry places, an other nature. Whereof he giveth a notable example, where he sheweth that an olde woman of Athens used a long time, to eate Hemlocke (which is a ranke poison) first a little quantitie, and afterwarde more, till at length she could eate so much without hurt as would presently poison another. . . . So that custome in processe of time may alter nature.” Finally, we may quote one last saying of Cogan’s, because of the good sense and wisdom displayed in the sentiment, as true to-day as when it was written more than three hundred years ago: “Neither is it good for any man that is in perfect health, to observe any custome in dyet precisely, as Arnoldus teacheth upon the same verses in these wordes: Every man should so order himselfe, that he might be able to suffer heate and cold, and all motions, and meats necessary,168 so as he might change the houres of sleeping and waking, and his dwelling and lodging without harme: which thing may be done if we be not too precise in keeping custome, but otherwise use things unwonted. Which sentence of Arnoldus agraeth verie well to that of Cornelius Celsus: He that is sound and in good health, and at libertie, should bind himselfe to no rules of dyet. To need neither Phisition or Chirurgion, he must use a diverse order of life, and be sometimes in the countrie, sometime in the towne, sometimes hunt, and sometime hawke. But some man may demand of me how this may agree with that saying of the scholar of Salernus ‘if you would be free from physicians, let these three be your physician, a cheerful mind, rest, and a moderate diet.’ Whereunto I answer, that a moderate dyet is alwaies good, but not a precise dyet: for a moderate diet is, as Terence speaketh in Andria: To take nothing too much: which alwaies is to be observed. But if a man accustome himselfe to such meats and drinks as at length will breed some inconvenience in his bodie, or to sleepe or to watch, or any other thing concerning the order of his life, such custome must naedes be amended and changed, yet with good discretion, and not upon the sudden: because sudden changes bring harme and weaknesse, as Hippocrates teacheth. He therefore that will alter any custome in dyet rightly, must do it with three conditions, which are expressed by Hippocrates. Change is profitable, if it be rightly used, that is, if it be done in the time of health, and at leisure, and not upon the sudden.”
This noteworthy book written by Cogan was preceded by the writings of Louis Cornaro, the Venetian, who forty years before had published the first edition of his celebrated book, “The Temperate Life,” and who was a most ardent advocate of the benefits to be derived by living temperately, especially in matters of diet. The simple diet which served for the169 nourishment of the oldest peoples of Syria, Greece, Egypt, and of the Romans when they were at the height of their prosperity and culture, was advocated by Cornaro as conducing to longevity, better health, and greater comfort of mind and body. Himself a striking example of the effects of a reasonable abstinence in diet (the last edition of his book having been written at the age of ninety-five), his teachings have continued to attract attention down to the present day; and although we have no values in grams or calories expressive of his average food consumption, it is quite evident that Cornaro lived a very abstemious life, eating little of the heavier articles of diet common to his time and country. It is perhaps not strictly physiological to refer to these cases, yet they have value as representing a sentiment, common to the centuries now passed, that benefit was to be derived by mankind from greater care in the taking of food; that prevalent customs and habits were leading the people into intemperate modes of life, and that these were surely tending toward the physical and mental deterioration of the nation. We may attach much or little weight to these conclusions, but there is a certain degree of significance in the views, current then as now, that dietetic customs and habits have no real connection with bodily requirements.
Passing down to our own times, we find physiologists, by the aid of scientific methods, studying the effects of smaller amounts of food (smaller than custom prescribes) on the condition of the body, thereby evincing a certain degree of skepticism concerning the dietary standards based on habit and usage. This has been especially true regarding the nitrogen requirement, or the need for proteid food. As has been clearly pointed out in other connections, there are two distinct needs which the body has for food; one for proteid or nitrogen, the other for energy-yielding material. According to the Voit standard, a man of average body-weight doing170 a moderate amount of work requires daily 118 grams of proteid food, or about 16 grams of metabolizable nitrogen, with fat and carbohydrate sufficient to yield a total fuel value of over 3000 large calories. As we have seen, the fuel value of the food must of necessity be a variable quantity because of variations in bodily activity. The more muscular work performed, the greater must be the intake of carbohydrate and fat, if the body is to be kept in equilibrium. With proteid or nitrogen, however, the case is quite different, since with adequate amounts of non-nitrogenous food, proteid is not drawn upon for the energy of muscular work. We can conceive of the nitrogen requirement, therefore, as being a constant factor in the well-nourished individual and dependent primarily upon body-weight, or more exactly, upon the weight of true proteid-containing tissue. Obviously, whatever else happens, there must be enough proteid food taken daily to maintain the body in nitrogen equilibrium. If this can be accomplished only by the ingestion of 16 grams of metabolizable nitrogen, then it is plain that the daily ration must contain at least 118 grams of proteid food; i. e., it must conform approximately at least to ordinary usage.
This question has been studied by many investigators, with very interesting and suggestive results. Thus, in 1887, Hirschfeld55 reported some experiments on himself, twenty-four years of age and weighing 73 kilos. His ordinary diet contained daily 100 to 130 grams of proteid, and the amount of nitrogen excreted varied from 16 to 20 grams per day, corresponding to a metabolism of proteid equal to the amount ingested. In other words, the body was essentially in nitrogen equilibrium. Then, for a period of fifteen days, during which he was unusually active, he lived on a diet in which the content of proteid corresponded to only 6 grams of nitrogen171 per day, and yet he remained in nitrogen equilibrium. The diet made use of was composed essentially of milk, eggs, rice, potatoes, bread, butter, sugar, and coffee, with some wine and beer, and on two days a little meat. It is to be observed that the nitrogen or proteid intake per day was only one-third of what he was accustomed to consume. In a second experiment, covering ten days, similar results were obtained. So that evidence was afforded that a young and vigorous man can maintain his body in nitrogen equilibrium, for fifteen consecutive days at least, on an amount of proteid food equal to only one-third of the minimal requirement called for by common usage. Plainly, the difference between a daily consumption of 118 grams of proteid food and 40 grams represents a large percentage saving, both of proteid and in the metabolism of proteid matter with all the attendant transformations. In these experiments, however, the subject consumed relatively large amounts of non-nitrogenous food, notably butter, of which on some days he took as much as 100 grams. The average fuel value of his food ranged from 3750 to 3916 calories per day; a fact of some importance, since it is to be remembered that both fat and carbohydrate tend to protect proteid metabolism.
In an experiment reported in 1889 by Carl Voit56, on a vegetarian weighing about 57 kilos, it was found that with a purely vegetable diet the subject was able, for a few days at least, to maintain his body in essentially a condition of nitrogen equilibrium on a daily diet containing 8.4 grams of nitrogen, corresponding to 52.5 grams of proteid. In addition, there was a large consumption of starchy food with some fat. Klemperer,57 experimenting with two young men,172 having a body-weight of 64 and 65.5 kilos, respectively, was able to keep them in a condition of nitrogenous equilibrium for a period of eight days on 4.38 grams and 3.58 grams of nitrogen per day. The diet, however, had a large fuel value, 5020 calories per day, and contained in addition to the small amount of proteid, 264 grams of fat, 470 grams of carbohydrate, and 172 grams of alcohol. Breisacher,58 in an experiment on himself, using a mixed diet composed of 67.8 grams of proteid, 494.2 grams of carbohydrate, and 60.5 grams of fat per day, with a total fuel value of 2866 calories, observed a daily excretion of nitrogen during thirty days of 8.23 grams. This corresponds to a metabolism of 51.4 grams of proteid, thus showing that the 67 grams of food-proteid taken was quite sufficient to maintain nitrogen equilibrium for the above length of time.
Caspari and Glässner59 have reported observations made on two vegetarians, a man and his wife, aged 49 and 48 years respectively, who had lived for some years exclusively on a vegetable diet. The man had a body-weight of 68.8 kilos, while the woman weighed 58 kilos. During five days, the man consumed per day, on an average, 7.83 grams of nitrogen and 4559 calories. This corresponds to 0.114 gram of nitrogen per kilo of body-weight, and 66 calories per kilo. On this diet, the man gained slightly in weight and showed a plus nitrogen balance of 5.2 grams for the five days. In other words, even this low nitrogen or proteid intake was more than sufficient to meet the wants of his body. The wife, during the same period of time, consumed per day 5.33 grams of nitrogen and 2715 calories, corresponding to 0.092 gram of nitrogen per kilo of body-weight and 47 calories per kilo.173 On this diet, the woman gained 0.9 kilo in weight during the five days, and like the man, she showed a plus nitrogen balance of 2.45 grams for the entire period. The somewhat noted experiments of Sivén have been referred to in another connection, and it will suffice to recall the fact that he was able, with a body-weight of 60 kilos, to establish nitrogen equilibrium on 6.26 grams of nitrogen, and for a day or two on 4.5 grams of nitrogen, with a total fuel value of only 2444 calories in the day’s ration.
These few illustrations will serve to indicate that, so far as the maintenance of nitrogen equilibrium is concerned during short periods of time, there is no necessity for the consumption of proteid food in such amounts as common usage dictates. The high proteid intake called for by the “standard dietaries,” and the ordinary practices of mankind, is not needed to establish a condition of nitrogen equilibrium. It would seem, however, as if results of this nature, presented from time to time by various investigators, have been considered more in the light of scientific curiosities than as data having an important bearing on physiological processes. So strong has been the hold upon the medical and physiological mind of the necessity of high proteid that such figures as the above have merely excited comment, without weakening in any measure the prevalent conviction that health, strength, and the power to work necessitate a high rate of proteid exchange.
To one willing to accept the data as having possible significance there arises at once the question, How long can the body be maintained in nitrogen equilibrium on such relatively small quantities of proteid food? In other words, can experiments of this nature, extending over comparatively short periods of time, be safely accepted as a reliable means of measuring the proteid requirements of the body for indefinite periods? Suppose, says the critic, we grant that the174 body can maintain itself in nitrogen equilibrium for a week or two on a very small amount of proteid food, what proof have we that in the long run the body will be benefited thereby, or even able to exist in a condition of normal strength and vigor? In other words, is a low proteid diet, one that seems sufficient to maintain the body in nitrogen equilibrium, a wholly safe one to follow? May there not be other elements to be considered, aside from nitrogen equilibrium, which, if fully understood, would satisfactorily account for the customs of mankind, in which perhaps man’s instincts have been followed for the betterment of the race? It was with a view to learning more concerning these questions that five years ago the writer commenced systematic, experimental, work upon the nutrition of man, with special reference to his nitrogen requirements. The experiments and observations have been continued up to the present time, with many suggestive results, some of which will now be referred to.60
One group of subjects was composed of professional men, professors and instructors in the university, whose work was mainly mental rather than physical, though by no means excluding the latter. Of this group, two cases will be referred to with some regard for detail, since in no other way can so striking a picture be presented of the effects produced. The first subject weighed 65 kilos in the fall of 1902, and at that time was nearly 47 years of age. His dietetic habits were in accord with common practice, and his daily consumption of proteid food averaged close to 118 grams. With a clear recognition of the principle that the habits of a lifetime should not be too suddenly changed, a very gradual reduction in the total amount of food, and especially of proteid matter, was made. This finally resulted, with this particular175 subject, in the complete abolition of breakfast, with the exception of a small cup of coffee. A light lunch was taken at noontime, followed by a more substantial dinner at night. There was no change to a vegetable diet, but naturally any attempt to cut off largely the amount of proteid food necessarily results in a marked diminution in the quantity of animal food or meats. It is a somewhat singular though suggestive fact, that a change of this order gradually results in a stronger liking for simple foods, with their more delicate flavor, accompanied by a diminished desire for the heavier animal foods.
As the day’s ration was gradually reduced in amount, the body-weight began to fall off, until after some months it became stationary at 57 kilos, at which point it has remained practically constant for over three years. The sixteen pounds of weight lost was composed, mainly at least, of superfluous fat. For a period of nine months, from October, 1903, to the end of June, 1904, the amount of proteid material broken down in the body was determined each day. The average daily metabolism of nitrogen for the entire period of nearly nine months amounted to 5.69 grams. For the last two months, it averaged 5.4 grams per day. Analyses made from time to time since these figures were obtained show that the subject is still living at the same low level of nitrogen metabolism. In fact, the data available afford satisfactory proof that for a period covering over three years this particular person has subsisted on an amount of proteid food equal to a metabolism of not more than 5.8 grams of nitrogen per day. It may be asked why the subject should have continued such a low proteid diet after the nine months’ period was completed? In reply, it may be said that the new habit has taken a firm hold, and entirely supplanted the dietetic desires and cravings of the preceding years. Further, the improved condition of health, freedom176 from minor ailments that formerly caused inconvenience and discomfort, and the greater ability to work without fatigue, have all combined to place the new habit on a firm basis, from which there is no desire to change.
Consider for a moment what this lowered consumption of proteid food really amounts to, as compared with ordinary usage and the so-called dietary standards. The latter call for at least 118 grams of proteid or albuminous food daily, of which 105 grams should be absorbable, in order to maintain the body in a condition of nitrogen equilibrium, and in a state of physical vigor and general tone. This would mean a daily metabolism and excretion of at least 16 grams of nitrogen. Our subject, however, excreted per day, during nine months, only 5.69 grams of nitrogen, which means a metabolism of 35.6 grams of proteid; i. e., about one-third the amount ordinarily deemed necessary to meet man’s requirement for proteid food. But was our subject in nitrogen equilibrium on this small amount of proteid food? We answer yes, as the following balance period shows:
Output. |
|||
Nitrogen in Food. |
Nitrogen through Kidneys. |
Weight of Excre- ment (dry). |
|
March 20 |
6.989 grams. | 5.91 grams. | 3.6 grams. |
21 |
6.621 | 5.52 | . . |
22 |
6.082 | 5.94 | 12.0 |
23 |
6.793 | 5.61 | 18.5 |
24 |
5.057 | 4.31 | 23.0 |
25 |
6.966 | 5.39 | 16.9 |
| ──── | ─── | ─── | |
| 74.0 grams contain 6.42% N. |
|||
| 38.508 | 32.68 + | 4.75 grams nitrogen. | |
| ───── | ────────── | ─── | |
| 38.508 grams nitrogen. 37.43 grams nitrogen. | |||
Nitrogen balance for six days = +1.078 grams. |
|||
Nitrogen balance per day = +0.179 gram. |
|||
Output. |
|||
Nitrogen in Food. |
Nitrogen through Kidneys. |
Weight of Excre- ment (dry). |
|
March 20 |
6.989 grams. | 5.91 grams. | 3.6 grams. |
21 |
6.621 | 5.52 | . . |
22 |
6.082 | 5.94 | 12.0 |
23 |
6.793 | 5.61 | 18.5 |
24 |
5.057 | 4.31 | 23.0 |
25 |
6.966 | 5.39 | 16.9 |
| ――― | ―― | ―― | |
| 74.0 grams contain 6.42% N. |
|||
| 38.508 | 32.68 + | 4.75 grams nitrogen. | |
| ――― | ――――――― | ―― | |
| 38.508 grams nitrogen. 37.43 grams nitrogen. | |||
Nitrogen balance for six days = +1.078 grams. |
|||
Nitrogen balance per day = +0.179 gram. |
|||
In this particular period of six days, the body was really gaining a little nitrogen, i. e., storing away a small amount of proteid for future use, although it may be granted that the amount was too small to have any special significance. During this period, the average daily intake of nitrogen was 6.4 grams, equal to 40 grams of proteid food. The average daily output of nitrogen through kidneys and excrement was 6.24 grams. The average daily output of metabolized nitrogen, through the kidneys, was 5.44 grams, corresponding to the breaking down of 34 grams of proteid material. Further, it should be stated that the total calorific value of the daily food during this period was less than 2000 calories. Let me add now a final balance period taken at the close of the nine months’ trial:
Output. |
|||
Nitrogen in Food. |
Nitrogen through Kidneys. |
Weight of Excre- ment (dry). |
|
June 23 |
6.622 grams. | 5.26 grams. | 10.6 grams. |
24 |
6.331 | 5.30 | 30.7 |
25 |
4.941 | 4.43 | 14.2 |
26 |
5.922 | 4.66 | 11.9 |
27 |
5.486 | 4.98 | 15.2 |
| ──── | ─── | ─── | |
| 82.6 grams contain 6.08% N. |
|||
| 29.302 | 24.63 + | 5.022 grams nitrogen. | |
| ───── | ────────── | ──── | |
| 29.302 grams nitrogen. 29.562 grams nitrogen. | |||
Nitrogen balance for five days = -0.350 gram. |
|||
Nitrogen balance per day = -0.070 gram. |
|||
Output. |
|||
Nitrogen in Food. |
Nitrogen through Kidneys. |
Weight of Excre- ment (dry). |
|
June 23 |
6.622 grams. | 5.26 grams. | 10.6 grams. |
24 |
6.331 | 5.30 | 30.7 |
25 |
4.941 | 4.43 | 14.2 |
26 |
5.922 | 4.66 | 11.9 |
27 |
5.486 | 4.98 | 15.2 |
| ――― | ―― | ―― | |
| 82.6 grams contain 6.08% N. |
|||
| 29.302 | 24.63 + | 5.022 grams nitrogen. | |
| ――― | ――――――― | ―― | |
| 29.302 grams nitrogen. 29.562 grams nitrogen. | |||
Nitrogen balance for five days = -0.350 gram. |
|||
Nitrogen balance per day = -0.070 gram. |
|||
In this period of five days, the average daily intake of nitrogen was 5.86 grams, corresponding to 36.6 grams of proteid food. The average daily output of metabolized nitrogen was 4.92 grams, implying the breaking down in the body178 of only 30.7 grams of proteid material per day. The fuel value of the daily food, calculated as closely as possible, was less than 2000 calories. The body was essentially in nitrogen equilibrium, the minus balance being too small to have any special significance.
It will be instructive to consider next the actual character and amount of the diet made use of on several of these balance days:
March 21.
Breakfast.—Coffee 119 grams, cream 30 grams, sugar 9 grams.
Lunch.—One shredded wheat biscuit 31 grams, cream 116 grams, wheat gem 33 grams, butter 7 grams, tea 185 grams, sugar 10 grams, cream cake 53 grams.
Dinner.—Pea soup 114 grams, lamb chop 24 grams, boiled sweet potato 47 grams, wheat gems 76 grams, butter 13 grams, cream cake 52 grams, coffee 61 grams, sugar 10 grams, cheese crackers 16 grams.
Total nitrogen content of the day’s food = 6.621 grams.
June 24.
Breakfast.—Coffee 96 grams, cream 32 grams, sugar 8 grams.
Lunch.—Creamed codfish 89 grams, baked potato 95 grams, butter 10 grams, hominy gems 58 grams, strawberries 86 grams, sugar 26 grams, ginger snaps 47 grams, water.
Dinner.—Cold tongue 14 grams, fried potato 48 grams, peas 60 grams, wheat gems 30 grams, butter 11 grams, lettuce-orange salad with mayonnaise dressing 155 grams, crackers 22 grams, cream cheese 14 grams, ginger snaps 22 grams, coffee 58 grams, sugar 10 grams.
Total nitrogen content of the day’s food = 6.331 grams.
June 25.
Breakfast.—Coffee 101 grams, cream 36 grams, sugar 13 grams.
Lunch.—Omelette 50 grams, bacon 9 grams, French fried potato 23 grams, biscuit 29 grams, butter 8 grams, ginger snaps 42 grams, cream cheese 17 grams, iced tea 150 grams, sugar 15 grams.
Dinner.—Wheat popovers 57 grams, butter 10 grams, lettuce-orange salad with mayonnaise dressing 147 grams, crackers 22 grams, cream cheese 21 grams, cottage pudding 82 grams, coffee 48 grams, sugar 11 grams.
Total nitrogen content of the day’s food = 4.941 grams.
179
June 27.
Breakfast.—Coffee 112 grams, cream 22 grams, sugar 10 grams.
Lunch.—Roast lamb 9 grams, baked potato 90 grams, wheat gems 47 grams, butter 12 grams, iced tea 250 grams, sugar 25 grams, vanilla éclair 47 grams.
Dinner.—Lamb chop 32 grams, creamed potato 107 grams, asparagus 49 grams, bread 35 grams, butter 17 grams, lettuce-orange salad with mayonnaise dressing 150 grams, crackers 21 grams, cream cheese 12 grams, coffee 63 grams, sugar 9 grams.
Total nitrogen content of the day’s food = 5.486 grams.
It can be seen that there was nothing especially peculiar in these dietaries, aside from their simplicity, except that the quantities were small. Meat was not excluded; there was no approach to a cereal diet; there were no fads involved, nothing but simple moderation in the amounts of nitrogen-containing foods. Further, there was perfect freedom of choice; full latitude to consider personal likes and dislikes in the selection of foods; anything that appealed to the appetite could be eaten, with the simple restriction that the amount taken must be small. During the balance days, naturally, every article of food had to be carefully weighed and analyzed, which fact undoubtedly tended to limit in some degree the variety of foods chosen, since increase in the number of articles meant increased labor in analysis. Quite noticeable, however, was the extreme constancy in the nitrogen-content of the daily diet, even on those days when the food was not weighed. In other words, there had been gradually acquired a new habit of food consumption, and the individual, unconsciously perhaps, rarely overstepped the limits fixed by the new level of proteid metabolism. This is a fact that has been conspicuous in nearly all of our experiments, where freedom of choice in the taking of food has been followed; and is in harmony with the view that after a lower level of proteid metabolism has once been established, and the body has become accustomed to the new conditions, there is little tendency180 for any marked deviation from the new standards of food consumption.
With maintenance of body-weight, together with nitrogen equilibrium through all these months; and with health, strength, and mental and physical vigor unimpaired, there is certainly ground for the belief that the real needs of the body were as fully met by the lowered consumption of proteid food as by the quantities called for by the customary standards. Finally, it should be noted that this particular subject was small in weight, and hence did not need so much proteid as a man of heavier body-weight would require. In recognizing this principle, we may for future comparison calculate the nitrogen requirement of the body, on the basis of the present results, per kilo of body-weight. With the weight of the subject placed at 57 kilos, and with an average daily excretion of nitrogen amounting to practically 5.7 grams, it is plain that this individual was quite able to maintain a condition of equilibrium with a metabolism of 0.1 gram of nitrogen per kilo of body-weight. Translated into terms of proteid matter, this would mean a utilization by the body of 0.625 gram of proteid daily per kilo of body-weight. Regarding the fuel value of the daily food, we need not be more precise than to emphasize the fact that so far as could be determined, on the basis of chemical composition, the heat value of the food rarely exceeded 1900 calories per day. If we make a liberal allowance, for the sake of precaution, it would seem quite safe to say that this particular individual, under the conditions of life and bodily activity prevailing, did not apparently need of fuel value more than 2000 calories per day, which would correspond to 35 calories per kilo of body-weight.
Let us turn now to the second subject in this group, a man of 76 kilos body-weight, 32 years of age, and of strong physique. His active life in the laboratory called for greater181 physical exertion than the former subject, and consequently there was need for greater consumption of non-nitrogenous food, with the accompanying increase in fuel value of the day’s ration. As in the preceding case, there was no prescribing of food, but a gradual and voluntary diminution of proteid material. During the last seven months and a half of the experiment, the average daily excretion of nitrogen through the kidneys amounted to 6.53 grams, equivalent to a metabolism of 40.8 grams of proteid matter daily; a little more than one-third the minimal quantity called for by common usage. At first, the body-weight of the subject gradually fell until it reached 70 kilos, at which point it remained fairly constant during the last five months. That the quantity of food taken was quite sufficient to maintain the body in a condition of nitrogen equilibrium is apparent from the results of a comparison of income and outgo of nitrogen, as shown in the following table:
Output. |
||||
Nitrogen in Food. |
Nitrogen through Kidneys. |
Weight of Excre- ment (dry). |
||
May 18 |
8.668 grams. | 6.06 grams. | 14 grams. | |
19 |
6.474 | 7.17 | 39 | |
20 |
6.691 | 6.33 | 30 | |
| ── | ||||
21 |
8.345 | 6.78 | 83 contain 6.06% N. = 5.03 grm. N. | |
22 |
7.015 | 5.70 | . . | |
23 |
9.726 | 5.75 | 38 | |
24 |
10.424 | 6.39 | 57 | |
| ──── | ─── | ── | ||
| 95 contain 5.76% N. = 5.47 grm. N. | ||||
| ─── | ||||
| 10.50 grm. N. | ||||
| 57.343 | 44.18 + 10.50 grams nitrogen. | |||
| ──── | ──────────── | |||
| 57.343 grams N. 54.68 grams nitrogen. | ||||
Nitrogen balance for five days = +2.663 grams. |
||||
Nitrogen balance per day = +0.380 gram. |
||||
Output. |
||||
Nitrogen in Food. |
Nitrogen through Kidneys. |
Weight of Excre- ment (dry). |
||
May 18 |
8.668 grams. | 6.06 grams. | 14 grams. | |
19 |
6.474 | 7.17 | 39 | |
20 |
6.691 | 6.33 | 30 | |
| ― | ||||
21 |
8.345 | 6.78 | 83 contain 6.06% N. = 5.03 grm. N. | |
22 |
7.015 | 5.70 | . . | |
23 |
9.726 | 5.75 | 38 | |
24 |
10.424 | 6.39 | 57 | |
| ――― | ―― | ― | ||
| 95 contain 5.76% N. = 5.47 grm. N. | ||||
| ―― | ||||
10.50 grm. N. |
||||
| 57.343 | 44.18 + 10.50 grams nitrogen. | |||
| ――― | ――――――――― | |||
| 57.343 grams N. 54.68 grams nitrogen. | ||||
Nitrogen balance for five days = +2.663 grams. |
||||
Nitrogen balance per day = +0.380 gram. |
||||
182
The average daily intake of nitrogen was 8.192 grams, equivalent to 51.2 grams of proteid food. The average amount of nitrogen excreted through the kidneys each day was 6.31 grams, corresponding to a metabolism of 39.43 grams of proteid matter. The plus balance of 0.380 gram of nitrogen per day shows that not only was the amount of proteid food consumed quite adequate to meet the demands of the body, but the latter was able to store up 2.3 grams of proteid per day. Regarding the character of the food taken by this subject, it should be stated that there was gradually developed a tendency toward a pure vegetarian diet. During the last seven months of the experiment, meats were almost entirely excluded. The diet voluntarily selected thus differed decidedly from that of the preceding subject in that it was much more bulky, contained a larger proportion of undigestible vegetable matter, and was richer in fats and carbohydrates, with a corresponding increase in fuel value. The exact character of the daily dietary is indicated by the following data of food consumption, on four of the days of the above balance period:
May 19.
Breakfast.—Banana 102 grams, wheat rolls 50 grams, coffee 150 grams, cream 50 grams, sugar 21 grams.
Lunch.—Omelette 20 grams, bread 57 grams, hominy 137 grams, syrup 68 grams, potatoes 128 grams, coffee 100 grams, cream 50 grams, sugar 21 grams.
Dinner.—Tomato purée 200 grams, bread 24 grams, fried sweet potato 100 grams, spinach 70 grams, Indian meal 100 grams, syrup 25 grams, coffee 100 grams, cream 40 grams, sugar 21 grams.
Total nitrogen content of the day’s food = 6.474 grams.
May 20.
Breakfast.—Sliced orange 140 grams, coffee 100 grams, cream 30 grams, sugar 21 grams.
Lunch.—Lima beans 40 grams, mashed potato 250 grams, bread 28 grams, fried hominy 115 grams, syrup 48 grams, coffee 100 grams, cream 30 grams, sugar 21 grams.
183
Dinner.—Consommé 150 grams, string beans 140 grams, mashed potato 250 grams, rice croquette 93 grams, syrup 25 grams, cranberry jam 95 grams, bread 19 grams, coffee 100 grams, cream 30 grams, sugar 21 grams.
Total nitrogen content of the day’s food = 6.691 grams.
May 21.
Breakfast.—Banana 153 grams, coffee 150 grams, cream 30 grams, sugar 21 grams.
Lunch.—Potato croquette 229 grams, bread 25 grams, tomato 123 grams, Indian meal 109 grams, syrup 48 grams, coffee 100 grams, cream 20 grams, sugar 14 grams.
Dinner.—Bean soup 100 grams, bacon 5 grams, fried potato 200 grams, bread 31 grams, lettuce-orange salad 47 grams, prunes 137 grams, coffee 100 grams, cream 25 grams, sugar 21 grams, banana 255 grams.
Total nitrogen content of the day’s food = 8.345 grams.
May 23.
Breakfast.—Banana 229 grams, coffee 125 grams, cream 25 grams, sugar 21 grams.
Lunch.—Consommé 75 grams, scrambled egg 15 grams, bread 58 grams, apple sauce 125 grams, fried potato 170 grams, rice croquette 197 grams, syrup 68 grams, coffee 100 grams, cream 30 grams, sugar 21 grams.
Dinner.—Vegetable soup 100 grams, potato croquette 198 grams, bread 73 grams, bacon 7 grams, string beans 120 grams, water ice 77 grams, banana 270 grams, coffee 100 grams, cream 30 grams, sugar 14 grams.
Total nitrogen content of the day’s food = 9.726 grams.
While the critic might justly say that these dietaries lack variety and would not appeal to a fastidious taste, there is force in the illustration which they afford of a simple diet being quite adequate to meet the wants of the body. Further, it should be emphasized that there is no special virtue in any of these dietaries, aside from their simplicity and low content of nitrogen. They represent individual taste and selection. Any other form of diet would answer as well, provided there was not too large an intake of proteid, and provided further the fuel value of the day’s ration was sufficient to meet the requirements for heat and work. Again, it might be said that with this latter subject the daily consumption184 of proteid food was considerably larger than with the first subject. This is indeed true, but it must be remembered that the second subject had a body-weight of 70 kilos during the last seven months, while the first subject weighed only 57 kilos. Obviously, with this marked difference in the weight of living tissue there must be a corresponding difference in the extent of proteid katabolism, and consequently a difference in the demand for proteid food.
As we have seen, the smaller subject for a period of many months showed a proteid katabolism equal to 0.1 gram of nitrogen, per kilo of body-weight, daily. The second and larger subject, on a totally different diet, for seven months and a half, metabolized daily, on an average, 6.53 grams of nitrogen. Taking the weight of the body at 70 kilos, it is readily seen that the nitrogen metabolized daily per kilo of body-weight was 0.093 gram, almost identical with the rate of nitrogen exchange found with the first subject. It is certainly very suggestive that these two individuals with their marked difference in body-weight, under different degrees of physical activity, and living on different forms of diet, with only the one point in common of voluntary restriction in the amount of proteid food, until a new habit had been acquired and a new level of proteid metabolism attained, should have quite independently reached exactly the same level of nitrogen exchange per kilo of body-weight. And when it is remembered that this was attained by the daily consumption of not more than one-third to one-half the minimal amount of proteid food called for by the dietetic customs of mankind, and with maintenance of all the characteristics of good health through this comparatively long period of time, there certainly seems to be justification for the opinion that the consumption of proteid food, as practised by the people of the present generation, is far in excess of the needs of the body. Referring for a moment to the calorific value of the food185 used by the second subject, in the last balance period, it is to be noted that the heat value per day averaged 2448 calories, as estimated on the basis of the chemical composition of the food. This would amount to 34 calories per kilo. Whether this figure is strictly correct is immaterial; it is certainly sufficiently so to warrant the statement that the needs of the body were fully met by an intake of food below the standards set by usage, and that maintenance of nitrogen equilibrium on a greatly diminished consumption of proteid food is possible without increasing the intake of non-nitrogenous matter.
Finally, as affording additional evidence, we may refer to a third subject in this group, a man of 65 kilos body-weight, 26 years of age, who for a period of six consecutive months maintained body-weight, nitrogen equilibrium, and a general condition of good health, with a proteid metabolism equal to 7.81 grams of nitrogen per day. During the last two months of the experiment, the average excretion of nitrogen per day amounted to 6.68 grams, corresponding to a metabolism of 0.102 gram of nitrogen per kilo of body-weight. This figure, it will be noted, is practically identical with the values obtained with the preceding subjects, calculated to the same unit of weight. Further, this third subject did not reduce his nitrogen intake by an exclusion of meat, but made use of his ordinary diet gradually reduced in amount. His daily consumption of proteid food averaged 55 grams, or 8.83 grams of nitrogen, and on this amount of proteid, without increasing the intake of fats and carbohydrates, he was quite able to do his work with preservation of physiological equilibrium.
Views so radically different from those commonly accepted can be made to carry weight, only by the accumulation of supporting evidence obtained under widely different conditions of life, and by methods which will defy criticism. It might be argued, and with perhaps some justification, that while186 professional men, with freedom from muscular work, may be able to live without detriment on a relatively small amount of proteid food, such a conclusion would not be warranted for the great majority of mankind with their necessarily greater muscular activity. We are confronted at once with the oft-heard statement that the laboring man requires more proteid food; he has a more vigorous appetite, and he must take an abundance of meat and other foods rich in proteid, if he is to maintain his ability as a worker. Note the statements already made in other connections regarding the food consumption of Maine lumbermen, of men on the football team, of trained athletes in general. These men consume large amounts of proteid daily, because their work demands it. If the demand did not really exist, they would not so agree in the use of high proteid standards, so runs the argument. The custom certainly does exist and is almost universally followed; men in training for athletic events deem it necessary to consume large amounts of proteid food. Custom and long experience sanction a high proteid diet, rich in nitrogen, for the development and maintenance of that strength and vigor that help to make the accomplished athlete. It is common knowledge to-day, however, that the energy of muscle work does not have its origin in the breaking down of proteid material, certainly not when there is an adequate amount of fat and carbohydrate in the diet. A high proteid intake must therefore be called for because of some subtle quality, not at present fully understood. It must not be subjected to criticism, however, because it is sanctioned by custom, habit, and common usage.
Still, I have ventured to experiment somewhat with a group of eight university athletes, all trained men, and with some surprising results. We have not space for details, but it may be mentioned that the men were young, from 22 to 27 years of age, and were experts in some field of athletic work.187 By a preliminary study of their ordinary dietetic habits, it was found that they were all large consumers of proteid food, with a corresponding high rate of proteid katabolism. One subject of 92 kilos body-weight, during ten days, showed an average daily excretion through the kidneys of 22.79 grams of nitrogen, implying a metabolism of 142 grams of proteid matter per day. On one of these days, the nitrogen excretion reached the high figure of 31.99 grams, corresponding to a metabolism of about 200 grams of proteid matter. Calculated per kilo of body-weight, this means a metabolism of 0.35 gram of nitrogen, or three and a half times the amount needed by the three professional men for the maintenance of nitrogen equilibrium. These subjects, with an intelligent comprehension of the point at issue, and with full freedom in the choice of food, gradually diminished their daily consumption of proteid material, at the same time cutting down very markedly the total consumption of food. The experiment extended through five months, and during the last two months, the average daily excretion of metabolized nitrogen of the eight men amounted to 8.81 grams per man. This corresponds to a metabolism of 55 grams of proteid matter.
Further, the average daily output of nitrogen through the kidneys during the preceding two months was in many cases nearly, if not quite, as low as during the last two months of the experiment. If we contrast this average daily exchange of 8.81 grams of nitrogen with the average output prior to the change in diet, it is easy to see that the men were living on about one-half the amount of proteid food they were formerly accustomed to take. Moreover, if the metabolized nitrogen for each individual, with one exception, is calculated per kilo of body-weight, it is seen to vary from 0.108 gram to 0.134 gram; somewhat higher than was observed with the older professional men, but not conspicuously so. Again, it is to be emphasized that the lowered intake of proteid food188 with these men was quite adequate to maintain their bodies in nitrogen equilibrium. We may cite a single case by way of illustration:
Output. |
|||
Nitrogen in Food. |
Nitrogen through Kidneys. |
Weight of Excre- ment (dry). |
|
May 18 |
8.119 grams. | 5.75 grams. | . . grams. |
19 |
9.482 | 6.64 | 15 |
20 |
10.560 | 8.45 | . . |
21 |
8.992 | 8.64 | . . |
22 |
9.025 | 8.53 | . . |
23 |
8.393 | 7.69 | 89 |
24 |
7.284 | 7.34 | 24 |
| ──── | ─── | ── | |
| 128 grams contain 6.40% N. |
|||
| 61.855 | 53.04 + | 8.192 grams nitrogen. | |
| ───── | ────────── | ──── | |
| 61.855 grams nitrogen. 61.232 grams nitrogen. | |||
Nitrogen balance for seven days = +0.623 gram. |
|||
Nitrogen balance per day = +0.089 gram. |
|||
Output. |
|||
Nitrogen in Food. |
Nitrogen through Kidneys. |
Weight of Excre- ment (dry). |
|
May 18 |
8.119 grams. | 5.75 grams. | . . grams. |
19 |
9.482 | 6.64 | 15 |
20 |
10.560 | 8.45 | . . |
21 |
8.992 | 8.64 | . . |
22 |
9.025 | 8.53 | . . |
23 |
8.393 | 7.69 | 89 |
24 |
7.284 | 7.34 | 24 |
| ――― | ―― | ― | |
| 128 grams contain 6.40% N. |
|||
| 61.855 | 53.04 + | 8.192 grams nitrogen. | |
| ――― | ――――――― | ――― | |
| 61.855 grams nitrogen. 61.232 grams nitrogen. | |||
Nitrogen balance for seven days = +0.623 gram. |
|||
Nitrogen balance per day = +0.089 gram. |
|||
The daily intake of nitrogen during this balance period averaged 8.83 grams, corresponding to 55.1 grams of proteid food. The metabolized nitrogen eliminated through the kidneys averaged 7.58 grams per day, thus showing a daily average metabolism of 47.37 grams of proteid matter. With a body-weight of 63 kilos, this individual was maintaining equilibrium on a metabolism of 0.120 gram of nitrogen per kilo of body-weight. The fuel value of the day’s food as estimated did not exceed 2800 calories, thus substantiating the general statement that there is no need for increasing the fuel value of the food in any attempt to maintain a lower nitrogen level. This particular individual, in his choice of food, unconsciously drifted—as he expressed it—toward a simple vegetable diet, without, however, excluding meat entirely.189 The following four dietaries will serve to illustrate the character and amount of his daily food:
May 21.
Breakfast.—Banana 106 grams, boiled Indian meal 150 grams, cream 50 grams, sugar 21 grams, bread 59 grams, butter 16 grams.
Lunch.—Lamb chop 37 grams, potato croquette 105 grams, tomato 216 grams, bread 55 grams, butter 13 grams, sugar 14 grams, water ice 143 grams.
Dinner.—Bean soup 100 grams, bacon 10 grams, fried egg 22 grams, fried potato 100 grams, lettuce salad 63 grams, coffee 100 grams, cream 50 grams, sugar 21 grams, stewed prunes 247 grams.
Total nitrogen content of the day’s food = 8.992 grams.
May 22.
Breakfast.—Orange 60 grams, oatmeal 207 grams, roll 46 grams, butter 14 grams, coffee 150 grams, cream 150 grams, sugar 35 grams.
Lunch.—Boiled potato 150 grams, boiled onions 145 grams, macaroni 130 grams, fried rice 138 grams, syrup 48 grams, ice cream 160 grams, cake 26 grams.
Dinner.—Celery soup 150 grams, spinach 100 grams, mashed potato 100 grams, bread 19 grams, coffee 100 grams, cream 50 grams, sugar 7 grams, strawberry short-cake 169 grams.
Total nitrogen content of the day’s food = 9.025 grams.
May 23.
Breakfast.—Sliced banana 201 grams, cream 100 grams, sugar 28 grams, griddle cakes 103 grams, syrup 48 grams.
Lunch.—Consommé 150 grams, rice croquette 140 grams, syrup 48 grams, fried potato 100 grams, bread 36 grams, butter 15 grams, apple sauce 90 grams, coffee 75 grams, sugar 7 grams.
Dinner.—Vegetable soup 100 grams, bacon 20 grams, potato croquette 50 grams, string beans 120 grams, macaroni 104 grams, bread 26 grams, water ice 184 grams.
Total nitrogen content of the day’s food = 8.393 grams.
May 24.
Breakfast.—Orange 80 grams, fried rice 186 grams, syrup 72 grams, coffee 100 grams, cream 50 grams, sugar 21 grams.
Lunch.—Celery soup 125 grams, bread 34 grams, butter 19 grams, boiled onion 127 grams, boiled potato 150 grams, tomato sauce 50 grams, stewed prunes 189 grams, cream 50 grams.
190
Dinner.—Tomato soup 125 grams, bread 21 grams, fried potato 100 grams, spinach 130 grams, cream pie 158 grams, coffee 100 grams, cream 50 grams, sugar 14 grams.
Evening.—Ginger ale 250 grams.
Total nitrogen content of the day’s food = 7.284 grams.
Here, again, we have dietaries not particularly attractive to every one, but they represent the choice of an individual who was following his own preferences, and like the preceding dietaries they are characterized by simplicity. In any event, they were quite adequate for the wants of the body, and their value to us lies in the proof they afford that a relatively small intake of proteid food will not only bring about and maintain nitrogen equilibrium for many months, and probably indefinitely, but that such a form of diet is equally as effective with vigorous athletes, accustomed to strenuous muscular effort, as with professional men of more sedentary habits. Further, these many months of observation with different individuals all lead to the opinion that there are no harmful results of any kind produced by a reduction in the amount of proteid food to a level commensurate with the actual needs of the body. Body-weight, health, physical strength, and muscular tone can all be maintained, in partial illustration of which may be offered two photographs of one of the eight athletes taken toward the end of the experiment; pictures which are certainly the antithesis of enfeebled muscular structure, or diminished physical vigor.

STAPLETON
Photograph taken in the middle of the experiment, in April
191
FURTHER EXPERIMENTS AND OBSERVATIONS BEARING ON TRUE FOOD REQUIREMENTS
Topics: Dietary experiments with a detail of soldiers from the United States army. General character of the army ration. Samples of the daily dietary adopted. Rate of nitrogen metabolism attained. Effect on body-weight. Nitrogen balance with lowered proteid consumption. Influence of low proteid on muscular strength of soldiers and athletes. Effect on fatigue. Effect on physical endurance. Fisher’s experiments on endurance. Dangers of underfeeding. Dietary observations on fruitarians. Observations on Japanese. Recent dietary changes in Japanese army and navy. Observations of Dr. Hunt on resistance of low proteid animals to poisons. Conclusions.
General acceptance of a new theory, or a new point of view, can be expected only when there is an adequate amount of scientific evidence on which the theory can safely rest. Facts cannot be ignored, and the larger the amount of supporting evidence the more certain becomes the general truth of the theory to which it points. Corroborative evidence, therefore, is always desirable, and he who would open up a new point of view must be zealous in accumulating facts to uphold his position. Critics there are without number who are ever ready to pick flaws in an argument or overturn a theory, especially if the one or the other stands opposed to their own point of view. This, however, is highly advantageous for the advance of sound knowledge, since it necessarily prompts the advocate to search in all directions for added data, by which he can build a bulwark192 of fact sufficient to defy just criticism. Further, the true scientific spirit demands persistent and painstaking effort in the search after truth, that error and misconception may be avoided.
In harmony with these ideas, our attempt to ascertain the real needs of the body for proteid food led us to enlarge our evidence by a series of experiments with still another body of men, i. e., a detail of soldiers from the United States army.61 This was a somewhat more difficult and ambitious undertaking, since the number of subjects involved was larger, and because with this group of men we could not expect quite that high degree of intelligent co-operation afforded by the preceding subjects. Still, this very fact was in a sense an added inducement, since it offered the opportunity of experimenting with a body of men who naturally would not take kindly to anything that looked like deprivation, and whose continued co-operation could be expected only by satisfying their natural demands for food. If this could be accomplished by an intelligent prescription in their daily diet, and the experiment brought to a successful conclusion, with maintenance of body-weight, nitrogen equilibrium, health, strength, and general vigor; with an intake of proteid food essentially equal to that adopted by the preceding subjects, corroborative evidence of the highest value would be obtained.
The detail was composed of a detachment of twenty men from the Hospital Corps of the army, under the command of a first lieutenant and assistant surgeon. They were located in a convenient house near to the laboratory, where they lived during their six months’ stay in New Haven, under military discipline, and subject to the constant surveillance of the193 commanding officer and the non-commissioned officers. Having well-trained cooks and assistants, with all necessary facilities for preparing and serving their food, with members of the laboratory staff to superintend the weighing of the food as it was placed before the men, and with intelligent clerks to attend to the many details connected with such an undertaking, a somewhat unique physiological experiment was started. Thirteen members of the detachment really took part in the experiment as subjects, and they represented a great variety of types: of different ages, nationalities, temperaments, and degrees of intelligence. They were men accustomed to living an active life under varying conditions, and they naturally had great liking for the pleasures of eating. Further, it should be remembered that, although the men had volunteered for the experiment, they had no personal interest whatever in the principles involved, and it could not be expected that they would willingly incommode themselves, or suffer any great amount of personal inconvenience. Again, there were necessary restrictions placed upon their movements, when relieved from duty, which constituted something of a hardship in the minds of many of the men and added to the irksomeness and monotony of their daily life. Regularity of life was insisted upon, and this was a condition which brought to some of the men a new experience. These facts are mentioned because their recital will help to make clear that, from the standpoint of the men, there were certain depressing influences connected with the experiment which would add to any personal discomfort caused by restriction of diet.
The ordinary army ration to which these men were accustomed was rich in proteid, especially in meat, and during the first few days they were allowed to follow their usual dietary habits, in order that data might be obtained bearing on their average food consumption. The details of one day’s food194 intake will suffice to show the average character and amount of the food eaten per man:
Breakfast.—Beefsteak 222 grams, gravy 68 grams, fried potatoes 234 grams, onions 34 grams, bread 144 grams, coffee 679 grams, sugar 18 grams.
Dinner.—Beef 171 grams, boiled potatoes 350 grams, onions 55 grams, bread 234 grams, coffee 916 grams, sugar 27 grams.
Supper.—Corned beef 195 grams, potatoes 170 grams, onions 21 grams, bread 158 grams, fruit jelly 107 grams, coffee 450 grams, sugar 21 grams.
It is not necessary to comment upon the large proportion of proteid matter in the day’s ration; the three large portions of meat testify clearly enough to that fact, while the three equally large volumes of coffee indicate a natural disposition toward generous consumption of anything available. Habit, reinforced by inclination, had evidently placed these men on a high plane of food consumption.
For a period of six months, a daily dietary was prescribed for the subjects; the food for each meal and for every man being of known composition, each article being carefully weighed, while the content of nitrogen in the day’s ration was so graded as to bring about a gradual reduction in the amount of proteid ingested. The rate of proteid katabolism was likewise determined each day by careful estimation of the excreted nitrogen, balance experiments being made from time to time in order to ascertain if the men were in a condition of nitrogen equilibrium. Finally, it should be mentioned that the subjects lived a fairly active life, having each day a certain amount of prescribed exercise in the university gymnasium, in addition to the regular drill and other duties associated with their usual work.
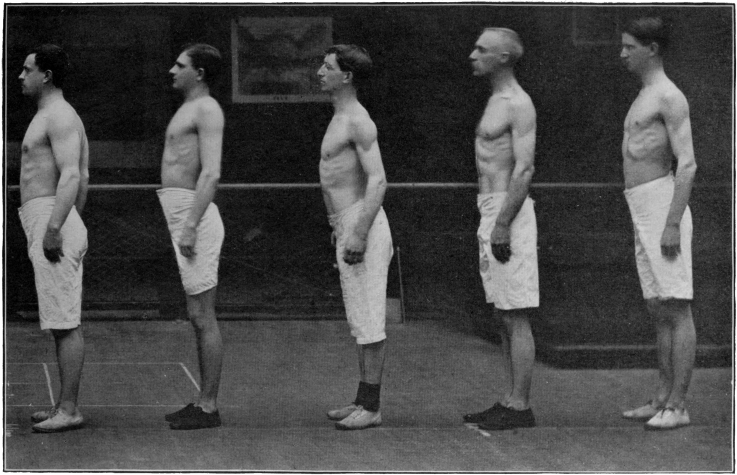
Photograph of the soldiers taken at the close of the experiment

Photograph of the soldiers taken at the close of the experiment
As just stated, the amount of proteid food was gradually reduced, three weeks being taken to bring the amount down to a level somewhat commensurate with the estimated needs of the body. This naturally resulted in diminishing largely the intake of meat, though by no means entirely excluding195 it. Effort was constantly made to introduce as much variety as was possible with simple foods, though the main problem with this group of men was to keep the volume of the food up to such a point as would dispel any notion that they were not having enough to eat. A second problem, which at first threatened trouble, was the fear of the men, as they saw the proportion of meat gradually drop off, that they were destined to lose their strength; but fortunately, they very soon began to realize that their fears in this direction were groundless, and a little later their personal experience opened their eyes to possible advantages which quickly drove away all further thought of danger, and made them quite content to continue the experiment. We may introduce here a few samples of the daily food given to the men after they had reached their lower level of proteid intake:
January 15.
Breakfast.—Wheat griddle cakes 200 grams, syrup 50 grams, one cup coffee62 350 grams.
Dinner.—Codfish balls (4 parts potato, 1 part fish, fried in pork fat) 150 grams, stewed tomato 200 grams, bread 75 grams, one cup coffee 350 grams, apple pie 95 grams.
Supper.—Apple fritters 200 grams, stewed prunes 125 grams, bread 50 grams, butter 15 grams, one cup tea 350 grams.
Total nitrogen content of the day’s food = 8.560 grams.
January 16.
Breakfast.—Soft oatmeal 150 grams, milk 100 grams, sugar 30 grams, bread 30 grams, butter 10 grams, one cup coffee 350 grams.
Dinner.—Baked macaroni with a little cheese 200 grams, stewed tomato 200 grams, bread 50 grams, tapioca-peach pudding 150 grams, one cup coffee 350 grams.
Supper.—Fried bacon 20 grams, French fried potato 100 grams, bread 75 grams, jam 75 grams, one cup tea 350 grams.
Total nitrogen content of the day’s food = 7.282 grams.
196
March 1.
Breakfast.—Fried rice 150 grams, syrup 50 grams, baked potato 150 grams, butter 10 grams, one cup coffee 350 grams.
Dinner.—Thick pea soup 250 grams, boiled onions 150 grams, boiled sweet potato 150 grams, bread 75 grams, butter 20 grams, one cup coffee 350 grams.
Supper.—Celery-lettuce-apple salad 120 grams, crackers 32 grams, American cheese 20 grams, potato chips 79 grams, one cup tea 350 grams, rice custard 100 grams.
Total nitrogen content of the day’s food = 7.825 grams.
March 3.
Breakfast.—Boiled hominy 175 grams, milk 125 grams, sugar 25 grams, baked potato 150 grams, butter 10 grams, one cup coffee 350 grams.
Dinner.—Hamburg steak with much bread, fat, and onions 150 grams, boiled potato 250 grams, bread 75 grams, butter 10 grams, one cup coffee 350 grams.
Supper.—Tapioca-peach pudding 250 grams, bread 75 grams, butter 20 grams, jam 75 grams, one cup tea 350 grams.
Total nitrogen content of the day’s food = 8.750 grams.
March 6.
Breakfast.—Sliced banana 100 grams, fried Indian meal 150 grams, syrup 50 grams, baked potato 150 grams, butter 10 grams, one cup coffee 350 grams.
Dinner.—Corned beef 50 grams, boiled cabbage 200 grams, mashed potato 250 grams, bread 75 grams, fried rice 100 grams, jam 75 grams, one cup coffee 350 grams.
Supper.—Crackers 32 grams, butter 10 grams, sardine 14 grams, sponge cake 150 grams, apple sauce 150 grams, one cup tea 350 grams.
Total nitrogen content of the day’s food = 10.265 grams.
March 30.
Breakfast.—Sliced banana 250 grams, fried hominy 150 grams, butter 10 grams, syrup 75 grams, one cup coffee 350 grams.
Dinner.—Codfish balls 125 grams, mashed potato 250 grams, stewed tomato 200 grams, bread 35 grams, apple sauce 200 grams, one cup coffee 350 grams.
Supper.—Chopped fresh cabbage with salt, pepper, and vinegar 75 grams, bread 50 grams, butter 20 grams, fried sweet potato 250 grams, cranberry sauce 200 grams, sponge cake 50 grams, one cup tea 350 grams.
Total nitrogen content of the day’s food = 9.356 grams.
197
March 31.
Breakfast.—Fried Indian meal 100 grams, syrup 75 grams, baked potato 250 grams, butter 20 grams, one cup coffee 350 grams.
Dinner.—Tomato soup, thick, with potatoes and onions boiled in, 300 grams, scrambled egg 50 grams, mashed potato 200 grams, bread 50 grams, butter 10 grams, one cup coffee 350 grams.
Supper.—Fried bacon 20 grams, boiled potato 200 grams, butter 10 grams, bread pudding 150 grams, sliced banana 200 grams, one cup tea 350 grams.
Total nitrogen content of the day’s food = 8.420 grams.
April 1.
Breakfast.—Fried hominy 150 grams, syrup 75 grams, baked potato 200 grams, butter 20 grams, one cup coffee 350 grams.
Dinner.—Baked spaghetti 200 grams, mashed potato 250 grams, boiled turnip 150 grams, bread 35 grams, butter 10 grams, apple sauce 200 grams, one cup coffee 350 grams.
Supper.—Fried bacon 25 grams, fried sweet potato 200 grams, bread 35 grams, butter 20 grams, jam 100 grams, apple-tapioca pudding 300 grams, one cup tea 350 grams.
Total nitrogen content of the day’s food = 7.342 grams.
These dietaries are fair samples of the daily food given the men during the last five months of the experiment. If we place the intake of nitrogen at 8.5 grams per day, or even 9 grams daily, it would mean at the most an average daily consumption of 56 grams of proteid; viz., about one-third the amount they were accustomed to take under their ordinary modes of life. Of greater interest, however, is the rate of proteid katabolism shown by these men under the above conditions of diet, during the five months’ period. The average daily output of metabolized nitrogen for each man ranged from 7.03 grams—the lowest—to 8.91 grams—the highest. An excretion of 7.03 grams of nitrogen per day means a katabolism, or breaking down, of 43.9 grams of proteid matter; while the excretion of 8.91 grams of nitrogen corresponds to a katabolism of 55.6 grams of proteid. The grand average, i. e., the average daily output of nitrogen198 of all the men for the five months’ period amounted to 7.8 grams per man, corresponding to an average daily katabolism of 48.75 grams of proteid. The heaviest man of the group had a body-weight of 74 kilograms, while his average daily output of metabolized nitrogen amounted to 7.84 grams. This corresponds to 0.106 gram of metabolized nitrogen per kilo of body-weight; a figure which agrees quite closely with the lowest figures obtained with the preceding subjects when calculated to the same unit of weight. Many of the men, however, metabolized considerably more nitrogen or proteid in proportion to their body-weight, due in a measure at least to the fact that they were being fed more liberally with proteid food than was really necessary for the needs of the body. In this group, we have a body of men doing a reasonable amount of physical work, who lived without discomfort for five consecutive months on a daily consumption of proteid food not much, if any, greater than one-third the amount called for by common usage, and the average fuel value of which certainly did not exceed 3000 calories per day. Indeed, so far as could be determined on the basis of chemical composition, the heat value of the food was quite a little less than this figure would imply.
If the relatively small amount of proteid food made use of in this trial was inadequate for the real necessities of the body, some indication of it would be expected to reveal itself, with at least some of the men, by the end of the period. One criticism frequently made is that the subject draws in some measure upon his store of body material. Should this be the case, it is evident that body-weight—in such a long experiment as this—will gradually but surely diminish. Further, the subject will show a minus nitrogen balance, i. e., there will be a constant tendency for the body to give off more nitrogen than it takes in. As bearing on the first point, the following table showing the body-weights of the men at the commencement199 of the experiment in October, and at the close of the experiment in April will be of interest:
TABLE OF BODY-WEIGHTS
October, 1903 |
April, 1904 |
|
|---|---|---|
kilos |
kilos |
|
| Steltz | 52.3 |
53.0 |
| Zooman | 54.0 |
55.0 |
| Coffman | 59.1 |
58.0 |
| Morris | 59.2 |
59.0 |
| Broyles | 59.4 |
61.0 |
| Loewenthal | 60.1 |
59.0 |
| Sliney | 61.3 |
60.6 |
| Cohn | 65.0 |
62.6 |
| Oakman | 66.7 |
62.1 |
| Henderson | 71.3 |
71.0 |
| Fritz | 76.0 |
72.6 |
| Bates | 72.7 |
64.3 (Feb.) |
| Davis | 59.3 |
57.2 (Jan.) |
As is readily seen, five of the men practically retained their weight or made a slight gain. Of the others, Coffman, Loewenthal, Sliney, and Cohn lost somewhat, but the amount was very small. Further, the loss occurred during the first few weeks of the experiment, after which their weight remained practically stationary. Fritz and Oakman lost weight somewhat more noticeably, but this loss likewise occurred during the earlier part of the trial. The accompanying photographs of Fritz, taken at the close of the experiment, show plainly that such loss of weight as he suffered did not detract from the appearance of his well-developed musculature. Certainly, the200 photographs do not show any signs of nitrogen starvation, or suggest the lack of any kind of food.
Of all the men, Bates was the only one who underwent any great loss of weight. He, however, was quite stout, and the work in the gymnasium, reinforced by the change in diet, brought about what was for him a very desirable loss of body-weight. It is evident, therefore, that there was no marked or prolonged loss of body-weight as a result of the continued use of the low proteid diet. Regarding the second point, viz., nitrogen equilibrium, the following illustrations will suffice to indicate the relationship existing between the income and outgo of nitrogen. A balance experiment with each of the men, lasting seven days, February 29 to March 6, is here shown, the figures given being the daily averages for the period:
Nitrogen of Food. |
Nitrogen of Urine. |
Nitrogen of Excrement. |
Nitrogen Balance. |
|
|---|---|---|---|---|
grams |
grams |
grams |
grams |
|
| Oakman | 9.52 |
7.24 |
1.76 |
+0.52 |
| Henderson | 9.40 |
7.90 |
1.00 |
+0.50 |
| Morris | 9.49 |
6.05 |
2.30 |
+1.14 |
| Coffman | 9.53 |
7.92 |
1.47 |
+0.14 |
| Steltz | 9.62 |
7.16 |
1.95 |
+0.51 |
| Loewenthal | 9.64 |
7.00 |
1.71 |
+0.95 |
| Cohn | 9.27 |
7.63 |
1.41 |
+0.23 |
| Zooman | 9.49 |
7.13 |
1.76 |
+0.60 |
| Sliney | 9.52 |
8.08 |
1.92 |
–0.48 |
| Broyles | 9.43 |
7.01 |
1.19 |
+1.23 |
| Fritz | 9.37 |
6.36 |
1.81 |
+1.20 |
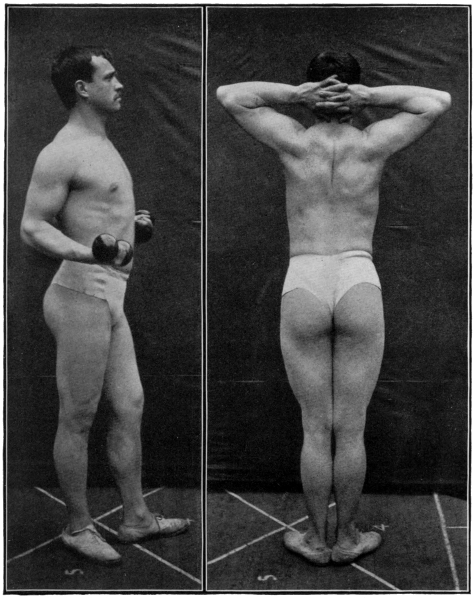
FRITZ
At the close of the experiment
With one exception, all of the men were plainly having more proteid food than was necessary to maintain the body201 in nitrogen equilibrium, the plus nitrogen balance in most cases being fairly large. It is only necessary to remember that a gain to the body of 1 gram of nitrogen means a laying by of 6.25 grams of proteid, and with such a gain per day it is apparent that the men were really being supplied with an excess of proteid food. This view is supported by the fact that a later balance experiment, when considerably less proteid food was being given, still showed many of the men in a condition of plus balance, or with a minus balance so small as to indicate essentially nitrogen equilibrium. The following figures, being daily averages of a balance period about the first of April, may be offered in evidence:
Nitrogen of Food. |
Nitrogen of Urine. |
Nitrogen of Excrement. |
Nitrogen Balance. |
|
|---|---|---|---|---|
grams |
grams |
grams |
grams |
|
| Broyles | 8.66 |
6.63 |
1.87 |
+0.16 |
| Fritz | 8.13 |
5.77 |
1.63 |
+0.73 |
| Loewenthal | 8.51 |
6.51 |
2.02 |
–0.02 |
| Steltz | 8.32 |
6.50 |
1.88 |
–0.06 |
| Cohn | 8.29 |
6.25 |
1.55 |
+0.49 |
| Morris | 8.45 |
6.49 |
2.27 |
–0.31 |
| Oakman | 8.62 |
7.04 |
1.87 |
–0.29 |
A daily intake of 8.5 grams of nitrogen means the consumption of 53 grams of proteid. Under these conditions of diet, the average daily amount of nitrogen metabolized was 6.45 grams, corresponding to 40.3 grams of proteid. The men were practically in a condition of nitrogen equilibrium, so that we are apparently justified in the general statement that the simple dietary followed with these men during the six months’ experiment, and which was accompanied by an average daily metabolism, after the first three weeks, of 7.8 grams of nitrogen, was certainly sufficient to maintain both body-weight202 and nitrogen equilibrium. Lastly, emphasis may be laid upon the fact that these values for nitrogen do not necessarily represent the minimal proteid requirement of the human body, since it is a well-established physiological principle that by increase of non-nitrogenous food the rate of proteid katabolism can always be further diminished; a principle which is plainly in harmony with the view that a high rate of proteid exchange is not a necessary requisite for the welfare of the body.
The experimental results presented afford very convincing proof that so far as body-weight and nitrogen equilibrium are concerned, the needs of the body are fully met by a consumption of proteid food far below the fixed dietary standards, and still further below the amounts called for by the recorded habits of mankind. General health is equally well maintained, and with suggestions of improvement that are frequently so marked as to challenge attention. Most conspicuous, however, though something that was entirely unlooked for, was the effect observed on the muscular strength of the various subjects. When the experiments were planned, it was deemed important to arrange for careful quantitative tests of the more conspicuous muscles of the body, with a view to measuring any loss of strength that might occur from the proposed reduction in proteid food. The thought that prompted this action was a result of the latent feeling that somehow muscular strength must be dependent more or less upon the proteid constituents of the muscles, and that consequently the cutting down of proteid food would inevitably be felt in some degree. The most that could be hoped for was that muscle tone and muscular strength might be maintained unimpaired. Hence, we were at first quite astonished at what was actually observed.
With the soldier detail, fifteen distinct strength tests were made with each man during the six months’ period, by203 means of appropriate dynamometer tests applied to the muscles of the back, legs, chest, upper arms, and forearms, reinforced by quarter-mile run, vault, and ladder tests, etc. The so-called “total strength” of the man was computed by multiplying the weight of the body by the number of times the subject was able to push up (strength of triceps muscles) and pull up (strength of biceps muscles) his body while upon the parallel bars, to this product being added the strength (dynamometer tests) of hands, legs, back, and chest. It should be added that all of these tests were made quite independently in the university gymnasium by the medical assistants and others in charge of the work there. It will suffice for our purpose to give here the strength tests of the various members of the soldier detail at the beginning and close of the experiment.
TOTAL STRENGTH
October. |
April. |
|
|---|---|---|
| Broyles | 2560 |
5530 |
| Coffman | 2835 |
6269 |
| Cohn | 2210 |
4002 |
| Fritz | 2504 |
5178 |
| Henderson | 2970 |
4598 |
| Loewenthal | 2463 |
5277 |
| Morris | 2543 |
4869 |
| Oakman | 3445 |
5055 |
| Sliney | 3245 |
5307 |
| Steltz | 2838 |
4581 |
| Zooman | 3070 |
5457 |
Without exception, we note with all of the men a phenomenal gain in strength, which demands explanation. Was it all due to the change in diet? Probably not, for these men at the beginning of the experiment were untrained, and it is not to be assumed that months of practical work in the gymnasium would not result in a certain amount of physical development, with corresponding gain in muscular skill and power. Putting this question aside for the moment, however, it is surely proper to emphasize this fact; viz., that although the men for a period of five months were restricted to a daily diet containing only one-third to one-half the amount of proteid food they had been accustomed to, there was no loss of physical strength; no indication of any physical deterioration that could be detected. In other words, the men were certainly not being weakened by the lowered intake of proteid food. This is in harmony with the principle, already discussed, that the energy of muscle work comes primarily from the breaking down of non-nitrogenous material, and consequently a diminished intake of proteid food can have no inhibitory effect, provided, of course, there is an adequate amount of proteid ingested to satisfy the endogenous requirements of the tissues.
On the other hand, recalling the large number of nitrogenous cleavage products which result from the breaking down of proteid material, we can conceive of an exaggerated exogenous proteid katabolism which may flood the tissues and the surrounding lymph with a variety of nitrogenous waste products, having an inhibitory effect upon the muscle fibres themselves, or upon the peripheral endings of the motor nerves, by which the muscles are prevented, directly or indirectly, from working at their highest degree of efficiency. This being true, a reduction of the exogenous katabolism to a level more nearly commensurate with the real needs of the body might result in a marked increase in the functional power of the tissue. However205 this may be, the fact remains that all of the subjects showed this great gain in strength; and furthermore, there was a noticeable gain in self-reliance and courage in their athletic work, both of which are likewise indicative of an improved condition of the body. How far these improvements are attributable to training and to the more regular life the men were leading, and how far to the change in diet, cannot be definitely determined. We may venture the opinion, however, for reasons to be made clear shortly, that the change in diet was in a measure at least responsible for the increased efficiency. As the writer has already expressed it, there must be enough food to make good the daily waste of tissue, enough food to furnish the energy of muscular contraction, but any surplus over and above what is necessary to supply these needs is not only a waste, but may prove an incubus, retarding the smooth working of the machinery and detracting from the power of the organism to do its best work.
Let us now turn our attention for a moment to the group of university athletes, remembering that these men had been in training for many months, and some of them for several years, prior to the commencement of the trial with a reduced proteid intake. In the words of the director of the gymnasium, “These eight men were in constant practice and in the pink of condition; they were in ‘training form’ when they began the changed diet.” Some of them had gained marked distinction for their athletic work; one during the early months of the test won the Collegiate and All-around Inter-collegiate Championship of America. Compare now the strength tests of these men as taken at the beginning and end of the five months’ experiment, during which they reduced their daily intake of proteid food more than fifty per cent:
206
TOTAL STRENGTH
January. |
June. |
|
|---|---|---|
| G. W. Anderson | 4913 |
5722 |
| W. L. Anderson | 6016 |
9472 |
| Bellis | 5993 |
8165 |
| Callahan | 2154 |
3983 |
| Donahue | 4584 |
5917 |
| Jacobus | 4548 |
5667 |
| Schenker | 5728 |
7135 |
| Stapleton | 5351 |
6833 |
It is to be observed that the majority of these trained men showed at the first trial in January a total strength test approximately equal to that of the soldier detail at the close of their experiment. This by no means implies that the latter men owed their gain in strength wholly to the systematic training they had undergone, but it is certainly plausible to assume that in a measure this was the case. In any event, it is plain that the long-continued low proteid diet of the soldiers had not interfered with a progressive muscular development, and the attainment of a high degree of muscular strength.
The noticeable feature in the figures obtained with the athletes, however, is the striking difference between the January and June results. Every man, without exception, showed a decided gain in his muscular power as measured by the strength tests. This improvement, to be sure, was not so marked as with the soldiers; a fact to be expected, since with these men the element of training and the acquisition of proficiency in athletic work could have played no part in the observed gain. Further, most of the tests indicated that the gain was progressive,207 each month showing an improvement, in harmony with the growing effect of the diminished proteid intake. With these subjects, the only tangible change in their mode of life which could in any sense be considered as responsible for their gain in strength was the change in diet. Consequently, it seems perfectly justifiable to conclude that the observations presented afford reasonable proof of the beneficial effects of a lowered proteid intake upon the muscular strength of man.
The significance of such a conclusion is manifestly obvious. It confirms and gives added force to the observations that man can profitably maintain nitrogen equilibrium, and body-weight, upon a much smaller amount of proteid food than he is accustomed to consume. It harmonizes with the view that the normal requirements of the body for food, under which health, strength, and maximum efficiency are best maintained, are on a far lower level than the ordinary practices of mankind would lead one to believe. The widespread opinion that a rich proteid diet, with the correspondingly high rate of proteid metabolism, is a necessity for the preservation of bodily strength and vigor, is seen to be without foundation; for even the most conservative estimate of the real value of these strength tests must carry with it the conviction that lowering the consumption of proteid food does not at least result in any weakening of the body. This is a fact of vital importance, for it needs no argument to convince even the most optimistic that while it might be possible to maintain body-weight and nitrogen equilibrium on a small amount of proteid food, such a form of physiological economy would not only be of no advantage to the individual, but would be positively injurious if there was a gradual weakening of the muscles of the body with decrease of physical strength, vigor, and endurance.
Another fact to be emphasized in this connection was the conviction, gradually acquired by many of the subjects, that208 they suffered less from fatigue after vigorous muscular effort than formerly. This was especially conspicuous in the case of Donahue, whose work on the Varsity basket-ball team called for vigorous exercise. It is interesting to note that this athlete, of 63 kilos body-weight, for the last four months of the experiment showed an average daily katabolism of 7.45 grams of nitrogen, corresponding to a breaking down of 46.5 grams of proteid material daily. Yet, with this low rate of proteid exchange, he maintained his position on the team with satisfaction to all, and with the consciousness of improved physical condition and greater freedom from fatigue. Other subjects, as the laboratory workers of the professional group, observed that the customary late afternoon fatigue, coincident with the continued walking and standing about the laboratory, gradually became far less conspicuous than usual; so that there seemed to be a consensus of opinion that in some way the change in diet was conducive to greater freedom from muscular weariness.
It is well understood by physiologists that the ability of a muscle to do work is inhibited by any condition that tends to depress the general nutritive state of the body, or that interferes with the local nutrition of the muscle or muscles involved. On the other hand, there are certain well-recognized conditions that tend to augment the power of the muscle, notably an increased circulation of blood through the tissue, the taking of food, and especially the introduction of sugar. Further, experiments have shown that when a given set of muscles has been made to work excessively, other muscles of the body quite remote will share in the fatigue, thus implying that muscular weariness and the diminished power to do work are connected with what may be termed fatigue products, which are distributed by means of the circulation. In this way, muscles and nerve endings alike are exposed to the inhibitory influence of waste products of unknown composition,209 formed in the muscle, and as previously stated, we may conceive of an exaggerated exogenous katabolism, with excessive proteid intake, by which muscular fatigue and weariness may be augmented; hence, the beneficial effect in this direction of a more rational food consumption, by which proteid katabolism shall be reduced to a true physiological level.
With these marked effects on strength and fatigue, it is reasonable to assume that some corresponding action may be exerted on physical endurance. As is well known, strength and endurance, though related, are quite distinct and can be separately measured. Strength tests, however, as usually carried out in gymnasium work, do involve in considerable degree the question of endurance, since it is customary to use as one of the factors in estimating total strength the number of times the man can pull up, or push up, his body on the parallel bars. Strictly speaking, however, the strength of a muscle is measured by the maximum force it can exert in a single contraction, while its endurance is estimated from the number of times it can contract well within the limit of its strength.
It is well known that endurance, both physical and mental, is one of the most variable of the human faculties, and it is usually considered that exercise or training is the chief cause of the differences so frequently seen. The Maine guide will row a boat or paddle a canoe for the entire day without undue fatigue, while the novice, though he may have the necessary strength, lacks the endurance to continue the task longer than a few hours. As expressed by Professor Fisher, “Some persons are tired by climbing a flight of stairs, whereas the Swiss guides, throughout the summer season, day after day spend the entire time in climbing the Matterhorn and other peaks; some persons are ‘winded’ by running a block for a street car, whereas a Chinese coolie will run for hours on end; in mental work, some persons are unable to210 apply themselves more than an hour at a time, whereas others, like Humboldt, can work almost continuously through eighteen hours of the day.” Again, Fisher states that “among some 75 tests of different persons holding their arms horizontal, many were found whose arms actually dropped against their will inside of 10 minutes, whereas several were able to hold them up over 1 hour, and one man held them 3 hours and 20 minutes, or a round 200 minutes, and then dropped them voluntarily. Similarly with deep knee-bending, some persons were found physically unable to rise again from the stooping posture after accomplishing less than 500 bendings, whereas several succeeded in stooping 1000 times, and in one case, 2400.” Here, we have inherent differences in endurance not associated with training or exercise, and the question may well be asked, What is the cause of these radical variations in the ability to repeat a simple muscular exertion?
Hitherto, little attention has been paid to the possible influence of diet upon this faculty. It has always been assumed that endurance, like physical strength, is augmented by a rich proteid diet, but it has never been considered that diet by itself was a factor of any great moment as compared with training or persistent exercise. It is true that claims have been advanced from time to time concerning the beneficial effects on endurance of a vegetable diet, and vegetarians have frequently presented glowing reports of the great increase in endurance they have experienced, but little attention has been given to such statements, and the matter has remained more or less in obscurity.
Recently, Professor Irving Fisher,63 of Yale, has conducted an interesting experiment on the influence of a change in diet on endurance, having the co-operation of nine healthy211 students as subjects. The experiment extended through five months, with endurance tests at the beginning, middle, and end of the period. At the outset, the men consumed daily an average of 2830 calories, of which 210 were in the form of flesh foods, such as meats, poultry, fish and shell-fish; 2.6 calories of proteid being ingested for each pound of body-weight. At the close of the experiment, the per capita calories had fallen to 2220, of which only 30 were in flesh foods, and the proteid had fallen to 1.4 calories per pound of body-weight. In other words, the total calories of the daily ration had dropped off about 25 per cent, the proteid about 40 per cent, and the flesh foods over 80 per cent, or to about one-sixth of their original amount.
To determine the endurance of the subjects, six simple gymnastic tests were employed, and one of mental endurance. The physical tests consisted of (1) in rising on the toes as often as possible; (2) deep knee-bending, or stooping as far as possible and rising to the standing posture, repeating as often as possible; (3) while lying on the back, raising the legs from the floor to a vertical position and lowering them again, repeating to the point of physical exhaustion; (4) raising a 5-lb. dumb-bell (with the triceps) in each hand from the shoulder up to the highest point above the head, repeating to the point of physical exhaustion; (5) holding the arms from the sides horizontally for as long a time as possible; (6) raising a dumb-bell (with the biceps) in one hand from a position in which the arm hangs free, to the shoulder and back, repeating to the point of physical exhaustion. This test was taken with four successive dumb-bells of decreasing weight, viz., 50, 25, 10, and 5 pounds respectively. The mental test consisted in adding specified columns of figures as rapidly as possible, the object being to find out whether the rapidity of performing such work tended to improve during the experiment.
212
The following table shows the results of the three sets of physical tests made in January, March, and June:
TESTS OF PHYSICAL ENDURANCE WITH THE NINE SUBJECTS
Time. |
B. |
E. |
Lq. |
Lw. |
M. |
P. |
R. |
T. |
W. |
|
|---|---|---|---|---|---|---|---|---|---|---|
| 1. Rising on | Jan. | 300 |
1007 |
333 |
69 |
127 |
1482 |
702 |
900 |
1263 |
| toes | Mar. | 400 |
1265 |
2620 |
65 |
400 |
831 |
1500 |
||
| June | 500 |
1061 |
3000 |
85 |
1500 |
1800 |
1263 |
1800 |
3350 |
|
| 2. Deep knee- | Jan. | 82 |
142 |
70 |
48 |
132 |
208 |
374 |
129 |
404 |
| bending | Mar. | 191 |
47 |
|||||||
| June | 200 |
81 |
202 |
58 |
155 |
230 |
453 |
250 |
508 |
|
| 3. Leg | Jan. | 25 |
52 |
9 |
22 |
30 |
27 |
50 |
23 |
30 |
| raising | Mar. | 33 |
34 |
40 |
||||||
| June | 33 |
38 |
20 |
35 |
31 |
37 |
103 |
19 |
53 |
|
| 4. 5lb. | Jan. | 75 |
138 |
78 |
38 |
51 |
44 |
100 |
83 |
185 |
| Dumb-bell | Mar. | 106 |
||||||||
| (triceps) | June | 127 |
59 |
80 |
51 |
75 |
56 |
104 |
101 |
501 |
m. s. |
m. s. |
m. s. |
m. s. |
m. s. |
m. s. |
m. s. |
m. s. |
m. s. |
||
| 5. Holding | Jan. | 5–0 |
1–33 |
4–7 |
3–37 |
3–30 |
5–39 |
2–5 |
3–22 |
11–0 |
| arms hori- | Mar. | 5–49 |
15–35 |
|||||||
| zontal | June | 9–36 |
2–56 |
3–50 |
3–0 |
6–5 |
10–1 |
3–16 |
3–24 |
23–45 |
| 6. 25lb. | Jan. | 50 |
18 |
16 |
6 |
20 |
11 |
10 |
25 |
54 |
| Dumb-bell | June | 105 |
10 |
26 |
33 |
30 |
29 |
27 |
75 |
108 |
| (biceps) |
The data presented show a marked improvement in March and June over the record made at the beginning of the experiment in January, except in the case of one subject, E. As Fisher states, the increased endurance observed can be ascribed only to dietetic causes, since no other factors of known significance could have aided in the result. The dietetic changes, as we have seen, consisted in a slight reduction of the total amount of food consumed daily, but with a large reduction of the proteid element, especially from flesh foods. It is significant, says Fisher, that the only man whose strength and endurance showed any decrease was E, “whose213 case was exceptional in almost all respects. His reduction in quantity of food, except for a spurt at the end, was less than of most of the men; his reduction in proteid, with the same exception, was the least of all; his reduction in quantity of flesh foods was the least of all.” He stands out conspicuously as the one man whose endurance failed to improve. The mental test carried out with the subjects pointed to “a slight increase in mental quickness,” but the adding test was too short to be of great value.
We see in these results another confirmation of the view that the welfare of the body is not impaired by a marked reduction in the amount of proteid food; on the contrary, benefit results in the increased efficiency which manifests itself in various directions. Physical endurance is an asset not to be ignored, and like the strength of an individual, it may well be fostered by the recognition and practice of a principle which seemingly has a firm physiological basis. Whether the fatigue poisons come from the excessive exogenous katabolism of proteids in general, or whether they are derived directly in a measure from flesh foods, need not be considered here; the main point is that by lowering the rate of proteid katabolism, which necessarily compels a reduction in the amount of flesh foods, there is a diminished quantity of nitrogenous waste floating about in the body. Further, we need not criticise too closely the method by which the reduction of food is accomplished; whether it be by encouraging mastication, with a view to better tasting and fuller enjoyment of the food, to the point of involuntary swallowing; or whether we follow natural taste and appetite, reinforced by the use of reason, with a full appreciation of the principle that the welfare of the body is best subserved by a quantity of food commensurate with true physiological needs.
In making this presentation of the true food requirements of the body as based on the results of physiological experimentation214 and observation, I am by no means unmindful of the dangers of underfeeding; but this is a condition comparatively rare. When occurring, as stated by Dr. Curtis, “it is either because of dyspepsia, in which case it really is involuntary, or comes from some silly notion born of a combination of innate mental crookedness and that ‘little knowledge’ that is a dangerous thing.” Overfeeding is the predominant dietetic sin, and with the prevailing dietary standards, as fixed by common usage, there is good ground for believing that it will continue for many years to come. Reason tells us, however, in the practice of our personal nutrition, to steer a middle course between physiological excess on the one side, and the minimal food requirement on the other. To quote again from Dr. Curtis,64 who has expressed the matter very forcibly, “The physiological chemist can easily draw a line on the Scylla (starvation) side of the channel. A dietary whereby the system gets less than it pays out is, obviously, a dangerous veer toward starvation rock. But on the Charybdis (stuffing) side, just as the whirlpool itself has no well-defined border, the channel boundary is not so easily marked. The case is exactly analogous to the stoking of a furnace. The proportion of ash to live coals is a telltale as to underfeeding, but not as to overfeeding. With undersupply of fuel the ashes overbalance the live coals, and the fire is thus foretold to be going out. But with an oversupply the fire simply burns the faster: all the fuel continues to be consumed; the more coal simply makes the more ash, so that equilibrium is not disturbed, although maintained at a higher level. To argue, therefore, that a given dietary is none too large, because the balance between the material receipts and expenditures of the economy is not upset, would be like saying that a given215 furnace-fire is certainly none too hot, since the ashes raked out of the fire-box just correspond to the amount of coal shovelled in. The same would be equally true of a slower fire consuming much less fuel. The philosophy of the matter is, then, to find the minimum of steam that will run the engine, and then maintain a fire somewhat hotter than the exact requirement, in order to run no risk of failure; or, to return to the metaphor already employed, the would-be careful liver must simply note how close to Scylla other voyagers have sailed with safety, and then steer his own bark accordingly.”
As one looks through the many careful dietary studies that have been made in recent years, it is easy to find striking illustrations of people, and communities of people, who have lived for long periods of time on dietaries so strikingly simple and meagre that it seems difficult at first glance to believe their daily needs could have been entirely satisfied. Yet, such observations are quite in accord with the facts we have been presenting, and they afford additional evidence that the artificial dietary standards that have been set up are widely at variance with the real requirements of the body for food. It may be quite true that many of the people referred to have been and are faddists, with peculiar notions regarding food, based on religious or other scruples, but that has no bearing on the main contention that they have lived for many years on amounts of food ridiculously small as compared with the ordinary customs of mankind. Thus, in Professor Jaffa’s report65 of investigations made among fruitarians and Chinese of California is an interesting account of a dietary study of a family of fruitarians, consisting of two women and three children. They had all been fruitarians from five to seven years, their diet being limited to nuts and fruit, except for216 the addition of celery, honey, olive oil, and occasionally a small amount of prepared cereal food. This family was in the habit of taking only two meals a day; at 10.30 in the morning and at 5 o’clock in the afternoon. The first meal always consisted of nuts and fruit, the nuts being eaten first. At the second meal, nuts were usually replaced by olive oil and honey. The nuts made use of were almonds, Brazil nuts, pine nuts, pignolias (a variety of pine nuts), and walnuts. Fruits, both fresh and dried, were used, the former including apples, apricots, bananas, figs, grapes, olives (pickled), oranges, peaches, pears, plums, and tomatoes. The dried fruits were dates and raisins.
On this limited dietary of raw, uncooked food, with a complete absence of the high-proteid animal foods, and the ordinary vegetables, legumes, etc., and without eggs or milk, this family, with three growing children, had lived all these years. Note now what Jaffa observed regarding their food consumption. The first subject, a woman 33 years of age and weighing 90 pounds, was studied for twenty consecutive days, all the food eaten being carefully weighed and its chemical composition determined. As a result, it was found that the average amount of food consumed per day was: proteid, 33 grams; fat, 59 grams; carbohydrate, 150 grams; with a total fuel value of 1300 calories. The other members of the family were studied in a similar manner, one of the children being the subject on two separate occasions. The table (on page 217), showing the average daily food consumption, gives a summary of the results obtained.
Proteid. |
Fat. |
Carbo- hydrate. |
Fuel Value. |
Proteid per Kilo Body- weight. |
|
|---|---|---|---|---|---|
grams |
grams |
grams |
calories |
grams |
|
| Woman, 33 years old, Weight 90 lbs. (40.9 kilos) |
33 |
59 |
150 |
1300 |
0.80 |
| Woman, 30 years old, Weight 104 lbs. (47.3 kilos) |
25 |
57 |
90 |
1040 |
0.52 |
| Girl, 13 years old, Weight 75 12 lbs. (34.3 kilos) |
26 |
52 |
157 |
1235 |
0.75 |
| Boy, 9 years old, Weight 43 lbs. (19.5 kilos) |
27 |
56 |
152 |
1255 |
1.38 |
| Girl, 6 years old, Weight 30 12 lbs. (13.9 kilos) |
24 |
58 |
134 |
1190 |
1.72 |
| Girl, 7 years old, Weight 34 lbs. (15.4 kilos) |
40 |
72 |
134 |
1385 |
2.59 |
As Professor Jaffa states, the tentative dietary standard for a woman at light work calls for 90 grams of proteid daily, with a fuel value of 2500 calories. Both of these women were light in weight, and furthermore had no occasion to do much physical work; but even so, a daily consumption of only 0.8 gram and 0.52 gram of proteid, respectively, per kilo of217 body-weight, with the small calorific values indicated, represents a phenomenally small amount of food. And yet Jaffa, in referring to the woman with the lowest intake of food, states that even this small quantity of food, judging from the appearance and manner of the subject, “seemed sufficient for her needs, enabling her to do her customary housework and take care of her two nieces and nephew.” Regarding the children, it is stated that the commonly accepted American dietary standard for a child 13 years old and of an average activity calls for about 90 grams of proteid and 2450 calories. As is seen from the table, however, the 13-year-old girl consumed of proteid less than one-third, and of fuel value only about 60 per cent of the amount called for; yet, says Jaffa, “notwithstanding the facts brought out by this comparison, the subject had all the appearances of a well-fed child in excellent health and spirits.”
We need not consume time in discussing the details of this experimental study, though the facts are interesting and suggestive, for it is only the general question of proteid requirement and calorific value that has interest for us at present. The fact is perfectly clear that this family of fruitarians, young and old, were quite able to live and thrive on a218 diet, the value of which in proteid and calories was at as low a level as was attained in our experimental studies. The rock of starvation, however, was not touched or even sighted by the voyagers down this stream of nutrition. We may all agree that it would be preferable, as a rule, to acquire the proteids, fats, and carbohydrates of our diet from a greater variety of sources than did the fruitarians; we might well complain at a dietary so limited in quality; but the point to be emphasized is that the low intake of proteid and the low fuel value were quite adequate for meeting the needs of the body. “It is a difficult matter,” says Professor Jaffa, “to draw any general conclusions from the foregoing dietaries without being unjust to the subjects. It would appear, upon examining the recorded data and comparing the results with commonly accepted standards, that all the subjects were decidedly undernourished, even making allowances for their light weight. But when we consider that the two adults have lived upon this diet for seven years, and think they are in better health and capable of more work than they ever were before, we hesitate to pronounce judgment. The three children, though below the average in height and weight, had the appearance of health and strength. They ran and jumped and played all day like ordinary healthy children, and were said to be unusually free from colds and other complaints common to childhood.”
Turning now to a larger community,—the island nation of Japan,—whose exploits in war have recently attracted the attention of the civilized world, we find a people the great majority of whom have remained untouched by the prodigality of western civilization, and whose customs and habits still bear the imprint of simplicity and frugality. After the restoration of Japan and the reorganization of the government in 1867, much attention was directed to the methods of living and to the dietary habits of the people, with the result that during219 the last twenty-five years there have been slowly accumulating many important data bearing on the food consumption of the people. These have recently been brought together in an interesting volume by Kintaro Oshima, and published66 in the English language.
| Subjects. | Body- weight. |
Digestible Nutrients and Energy per Man per Day. |
|||
|---|---|---|---|---|---|
| Proteid. | Fat. |
Carbo- hydrate. |
Fuel Value. |
||
kilos |
grams |
grams |
grams |
calories |
|
| School business agent | 57.5 |
65.3 |
11.3 |
493.8 |
2467 |
| Physician | . . . . |
61.9 |
8.0 |
468.5 |
2315 |
| Merchant | 47.6 |
81.5 |
19.6 |
366.2 |
2082 |
| Medical student | 49.0 |
74.8 |
11.2 |
326.9 |
1811 |
| Medical student | 48.5 |
64.7 |
5.1 |
469.6 |
2305 |
| Military cadets | . . . . |
72.3 |
11.7 |
618.1 |
3021 |
| Prisoners without work | 47.6* |
36.3 |
5.6 |
360.4 |
1726 |
| Prisoners at light work | 48.0* |
43.1 |
6.2 |
443.9 |
2112 |
| Prisoners at hard work | . . . . |
56.7 |
7.5 |
610.8 |
2884 |
| Physician | 40.2 |
48.3 |
15.5 |
438.2 |
2201 |
| Hygienic assistant | 40.5 |
46.5 |
19.7 |
485.3 |
2430 |
| Medical student | 51.0 |
42.8 |
14.0 |
438.2 |
2163 |
| Police prisoners | . . . . |
42.7 |
8.7 |
387.3 |
1896 |
| Army surgeon | 54.0 |
79.3 |
11.7 |
502.0 |
2567 |
| Soldier | 66.7 |
75.8 |
13.5 |
563.8 |
2828 |
| Soldier | 61.0 |
58.8 |
11.3 |
467.8 |
2330 |
| Soldier | 56.7 |
55.2 |
10.9 |
459.6 |
2276 |
* Average weight of twenty subjects.
As is well known, the great majority of the people of Japan220 live mainly on a vegetable diet. It is also known to physiologists at least that Japanese dietaries are characterized by a relatively small amount of proteid, though since the passage of the Food Supply Act of the navy in 1884, the proteid-content of the navy ration has been decidedly increased. It will be interesting to note a few of the results collated by Oshima, and some of the conclusions that he draws from the data presented. The foregoing table shows a few of the more striking results of the dietary studies obtained with various classes of people, where the food used was largely vegetable, but generally with some admixture of fish or meat.
The figures presented, which represent the actual amounts of food consumed, with proper correction for the indigestible portion, show a much smaller intake of proteid than is common with European and American people; indeed, both proteid and fuel value are very much less than common practices call for among western peoples, even when due allowance is made for differences in body-weight. To quote from Oshima, “Probably the most interesting of the dietary studies are those with poorer classes, which comprise by far the larger part of the population. The dietaries of the miscellaneous class, including employees, prisoners, etc., consisted largely of vegetable foods and supplied on an average 59 grams of proteid and 2190 calories of energy per man per day.” Especially suggestive were the results of a study made with a military colonist, a type of man very common in Japan; in reality farmers who live at home, but have military drill at certain fixed times. The subject was carefully selected under advice of officers in charge of the district, and weighed 59.9 kilograms. His diet consisted solely of cereals and vegetables, being identical with that of the people in the rural districts of Japan. His daily food was found to be composed of 46.3 grams of digestible proteid, with a fuel value of 2703 calories.
Even more striking were the results obtained in a study of221 the dietary habits of three healthy natives of Formosa, employed as day laborers at the military hospital. They weighed respectively 60.9, 55, and 54.8 kilograms. The main portion of their diet was rice, supplemented, however, by a little salt fish, salted melon, spinach, ginger, and greens. The daily amount of proteid ingested was 48.0 grams (37.4 grams of digestible proteid), with a total fuel value of 1948 calories. A composite sample of urine covering seven days showed an average daily output of metabolized nitrogen of 6.93 grams, corresponding to a breaking down of 43.3 grams of proteid.
Especially interesting also is a series of experiments with professional men, reported by Oshima, in which attention was paid to nitrogen balance. The following table shows the essential results:
Subject. |
Body- weight. |
Character of Food. |
Digestible Nutrients and Energy per Man per Day. |
||||
|---|---|---|---|---|---|---|---|
Proteid. |
Fat. |
Carbo- hydrate. |
Fuel Value. |
Nitrogen Balance. |
|||
kilos |
grams |
grams |
grams |
calories |
|||
N. K. |
43.1 |
mixed diet |
72.7 |
18.3 |
380.7 |
2091 |
+ |
S. A. |
49.5 |
mixed diet |
69.8 |
20.2 |
410.7 |
2222 |
+ |
N. K. |
42.9 |
mixed diet |
64.4 |
8.5 |
396.3 |
2028 |
+ |
N. K. |
43.2 |
mixed diet |
62.8 |
8.7 |
433.2 |
2178 |
+ |
N. K. |
43.0 |
vegetable |
68.5 |
19.7 |
433.0 |
2303 |
+ |
N. K. |
43.9 |
vegetable |
36.8 |
6.6 |
381.0 |
1824 |
- |
N. K. |
42.4 |
vegetable |
40.5 |
8.7 |
462.6 |
2200 |
+ |
S. A. |
49.6 |
vegetable |
34.4 |
7.5 |
451.9 |
2119 |
- |
S. A. |
49.9 |
vegetable |
43.5 |
9.1 |
500.0 |
2376 |
+ |
It is to be observed that in all of the above experiments, excepting two, the subjects gained nitrogen even with the low proteid intake and the small fuel value of the day’s food. Particularly noteworthy, in harmony with previous statements,222 are the results of the sixth and seventh experiments. In the sixth experiment, the subject was not able to maintain nitrogen equilibrium on a diet containing 36.8 grams of digestible proteid and having a fuel value of 1825 calories, but by raising the intake of carbohydrate food (seventh experiment) to 462 grams daily, thereby increasing the fuel value of the daily ration to 2200 calories (with a slight increase in the proteid incidental thereto), the body was able to change its previous loss of nitrogen into a gain; in other words, the added carbohydrate served as a protector of proteid.
The series of experiments as a whole, however, is to be considered in the light of additional data bearing on the dietary customs of a people who for generations have apparently lived and thrived on a daily ration noticeably low in its content of proteid, as well as low in its calorific value. As Oshima states, “It is probably fair to infer that the amount of proteid in the dietaries of the classes living largely on vegetable foods (and they constitute the larger part of the population) may not be very far from 60 grams per day,” or 45 grams of digestible proteid. It is reasonable to assume that the people live in this way from force of habit or of necessity, and we may agree with Baelz, a professor connected with the medical faculty of Tokyo University, “that their diet is sufficient from a physiological standpoint.” Doubtless a mixed diet, with a larger proportion of animal food, did their means readily permit, would offer some advantages from the standpoint of palatability and variety, but it is questionable if any material gain in health or strength would result. “It is sometimes remarked,” says Oshima, “that the peasants in the rural districts of Japan, living largely on vegetable food, are really healthier and stronger than people of the better classes, who live on a mixed diet, and the better physical condition of the former is commonly believed to be due to their diet.” This, however, is a difficult matter to decide,223 since there are so many other factors that are liable to play a part, such as the general conditions of life which are so widely different in the two classes.
It is plainly evident that the daily diet of the great bulk of the Japanese people has been characterized by a very low proteid standard, as contrasted with the standards and usages of the majority of European and American people. The fact is brought forward merely as confirmatory evidence, on a large scale, of the perfect safety of lowering the consumption of proteid food to somewhere near the level of the physiological requirements of the body. Generations of low proteid feeding, with the temperance and simplicity in dietary matters thereby implied, have certainly not stood in the way of phenomenal development and advancement when the gateway was opened for the ingress of modern ideas from western civilization. Many changes are sure to follow in the footsteps of the nation’s progress, and among these it is safe to prophesy that as public and private wealth, and resources in general, increase, the dietary of the people will gradually assume a more varied character with corresponding increase in volume. Whether such a change will prove of real benefit to the race, time alone can determine.
Having said so much concerning the Japanese, it is proper that a few additional statements should be made. The stature and general physique of the people could be advantageously improved. Is this a question of dietary, or is it connected with some condition of life on which the daily food has no bearing; or is it, perchance, a racial characteristic so deeply ingrained that conditions of environment are without noticeable influence? These questions cannot be definitely answered at present. Finally, we may call attention to the dietary changes inaugurated in recent years in connection with the new organization of the imperial army and navy. With a view to increasing the efficiency of the men, following224 the customs of other countries, an act was passed increasing the amount of proteid food in the navy dietary. Oshima’s report of the various steps taken to accomplish this end, with the results that followed, is interesting in several ways.
“A large part of the rice was to be replaced by bread, and meats were to be used liberally. The experience, during the first year that this ration was tried, indicated that bread and meat could not be advantageously substituted immediately for the rice, because most of the marines were unaccustomed to these food materials; consequently, a modification of the ration was introduced in 1885, whereby a rice-barley mixture was adopted in place of the bread. Barley was considered at that time as a better article of food than rice, on account of its higher proteid content, but later investigations showed that the digestibility of the nutrients of barley was small. In 1886, an effort was again made to substitute bread for the rice-barley mixture. In 1890, the ration allowance was reduced by one-fifth and an amount of money equivalent to the cost of the reduction in diet was given to each marine with which to buy accessory food according to his own choice. In 1898, the reduction was made one-tenth, instead of one-fifth as in previous years. In 1900, the cash allowance was abolished and a new ration adopted.” This ration contains about 150 grams of proteid (animal and vegetable food) and has a fuel value of over 3000 calories. In all of these changes, the proportion of rice was greatly reduced.
Probably, one of the chief reasons why persistent efforts were made to improve the dietary of the navy was the prevalence among the men of the disease known as beriberi. “While no satisfactory explanation as to the cause of the disease was offered, it was generally believed that there was some very close relation between the disease and the rice diet” (Oshima). During the years 1878–1883 inclusive, nearly 33 per cent of the marines suffered from beriberi. With the adoption of the new ration in 1884, in which a large part225 of the rice was replaced by bread and other articles, and with better hygienic conditions, this disease immediately began to disappear, and during the six years after the adoption of the new diet only 16 per cent of the marines were affected by the disease. Later on, hardly more than two or three cases a year were recorded. Advocates of a high proteid diet bring forward this illustration as an evidence of the danger connected with a lowered proteid intake; i. e., that the nutrition of the body will be impaired and diseases of various sorts liable to follow. Yet, Oshima is very careful to state, “It should be especially noted that here no attempt has been made to indicate the cause of beriberi or the relation between the disease and the diet.” That rice in itself can be a cause of the disease is not to be considered for a moment. Further, so far as any facts are concerned, the writer can see no ground for considering that a low rate of proteid metabolism has in itself any direct connection with the disease. From a dietary standpoint, it seems far more plausible to assume that the great restriction in variety of foods, so strikingly manifest in the dietary of the poorer people of Japan, results in a lack of some one or more elements which conduces to the disease, just as in scurvy the lack of fresh vegetables on long voyages was liable to be followed by an epidemic of this disease.
Consider the natural character of the dietary of the great bulk of the Japanese people, determined as it was by adverse financial circumstances. As Oshima states, “The rural population of the interior depends very largely or entirely upon a vegetable diet. Fish is eaten perhaps once or twice a month, and meat once or twice a year, if at all. The poorer working classes in the cities also use very little animal food. But the poorer classes in the city and the peasantry of the rural districts comprise nearly 75 per cent of the total population, and it is therefore safe to assume that this proportion lives226 chiefly, or wholly, upon vegetable diet. And this, it may be observed, means vegetarianism literally. The so-called lacto-vegetarianism is unknown in Japan. Cows are scarce, and milk and other dairy products are expensive, and such as are available are consumed almost entirely by the wealthier people in the cities.” It is also to be noted that the amount of fat in Japanese dietaries is very small. The reported data indicate that the usual vegetable dietaries contain only about 10 grams of fat per day, while even in the average mixed dietaries the amount rarely rises above 20 grams per day. In other words, the ordinary food of the Japanese was characterized by great lack of variety, and with such a preponderance of carbohydrate materials of a limited kind that it is easy to conceive of a possible dearth of some essential or accessory element, necessary for the preservation of that nutritive balance which aids in protection against disease.
If the resistance of the body to disease germs and toxic influences in general is really diminished by reducing the consumption of proteid food below the set dietary standards, then obviously here lies a tangible reason for the maintenance of a high proteid intake. I know of only one series of scientific observations that bears directly on this question. Dr. Reid Hunt of Washington has studied recently the power of resistance to the poison acetonitrile of animals kept for some time upon a reduced proteid diet. “My experiments,” says Dr. Hunt, “showed in all cases that the resistance was much increased.” In other words, the animals that had been fed a low proteid ration were able to endure a much larger dose of the poison than corresponding animals on their customary diet; “they resisted 2–3 times the ordinary fatal dose of acetonitrile.” This general subject, however, is obviously a very important one, and merits further experimental study under a diversity of conditions.
In conclusion, the facts here presented bearing on food requirements,227 especially those that relate to the need for proteid food, are seemingly harmonious in indicating that the physiological necessities of the body are fully met by a much more temperate use of food than is commonly practised. Dietary standards based on the habits and usages of prosperous communities are not in accord with the data furnished by exact physiological experimentation. Nitrogen equilibrium can be maintained on quantities of proteid food fully fifty per cent less than the every-day habits of mankind imply to be necessary, and this without increasing unduly the consumption of non-nitrogenous food. A daily metabolism of proteid matter equal to an exchange of 0.10–0.12 gram of nitrogen per kilogram of body-weight is quite adequate for physiological needs, provided a sufficient amount of non-nitrogenous foods—fats and carbohydrates—is taken to meet the energy requirements of the body.
The long-continued experiments on many individuals, representing different types and degrees of activity, all agree in indicating that equilibrium can be maintained indefinitely on these smaller quantities of food, and that health and strength can be equally well preserved, to say nothing of possible improvement. The lifelong experience of individuals and of communities affords sufficient corroborative evidence that there is perfect safety in a closer adherence to physiological needs in the nutrition of the body, and that these needs, so far as proteid food is concerned, are in harmony with the theory of an endogenous metabolism, or true tissue metabolism, in which the necessary proteid exchange is exceedingly limited in quantity. There are many suggestions of improvement in bodily health, of greater efficiency in working power, and of greater freedom from disease, in a system of dietetics which aims to meet the physiological needs of the body without undue waste of energy and unnecessary drain upon the functions of digestion, absorption, excretion, and metabolism228 in general; a system which recognizes that the smooth running of man’s bodily machinery calls for the exercise of reason and intelligence, and is not to be intrusted solely to the dictates of blind instinct or to the leadings of a capricious appetite.
229
THE EFFECT OF LOW PROTEID DIET ON HIGH PROTEID ANIMALS
Topics: A wide variety of foods quite consistent with temperance in diet. Safety of low proteid standards considered. Arguments based on the alleged effects of low proteid diet on high proteid animals. Experiments of Immanuel Munk with dogs. Experiments of Rosenheim. Experiments of Jägerroos. Comments on the above experiments. The experiments of Watson and Hunter on rats. The writer’s experiments with dogs. Details of the results obtained with six dogs. Comparison of the results with those of previous investigators. Effect of a purely vegetable diet on dogs. Different nutritive value of specific proteids considered. Possible influence of difference in chemical constitution of individual proteids. Effect of low proteid diet on the absorption and utilization of food materials in the intestine of dogs. General conclusions from the results of experiments with animals.
Man is by choice an omnivorous creature; he reaches out ordinarily in all directions for as wide a variety of foods as his circumstances and surroundings will allow. He rightly cultivates a taste for foods that have individuality of flavor, and derives pleasure and satisfaction from the eating of delicacies that appeal to palate and to reason. All this he can do without becoming an epicure or a glutton, and without violation of physiological laws or disregard of the teachings of temperance. As a being endowed with reason and intelligence he is, however, necessarily mindful of the possible deleterious effect of undue quantities of food, as he is likewise mindful of the desirability of avoiding certain varieties of food which personal experience has taught him are fraught with possible230 danger. Care and prudence in diet are legitimate outcomes of a reasonable interest in the welfare of the body, upon which so largely depend the happiness and working power of the individual.
The adoption of dietary habits that aim to accord with the physiological requirements of the body does not compel a crucifying of the flesh or a disregard of personal likes and dislikes. A reasonable intelligence combined with a disposition to exercise the same degree of judgment and care in the nutrition of the body as is expended on other matters, of no greater importance, pertaining to the individual, to the household, or to business interests, are all that is needed to bring about harmony between every-day dietary habits and the nutritive requirements of the body. There is no occasion, unless one finds pleasure and satisfaction in so doing, to resort to a limited dietary of nuts and fruits, to become an ardent disciple of vegetarianism, to adopt a cereal diet, to abjure meats entirely, or to follow in an intensive fashion any particular dietary hobby, except so far as may be necessary to insure an adequate amount of non-nitrogenous foods to meet the energy requirements of the body without unduly increasing the intake of proteid or nitrogenous food. Naturally, a man leading a life of great physical activity with the consequent demand for a large energy-yielding intake will be compelled to resort largely to vegetable foods, rich in starch and poor in proteid, or to eat largely of fatty foods. Reliance on meats and animal foods in general, under such conditions, would plainly involve a high proteid intake with a consequent high nitrogen metabolism, with the chance that even then the energy requirement would not be fully met.
In view of all that has been said, reinforced by the various facts brought forward as evidence, we must recognize the value of the non-nitrogenous foods as a source of energy, and this means plainly food from the plant kingdom. In any231 rational diet, vegetable foods of low nitrogen-content must predominate, while animal foods with their higher nitrogen values must be greatly subordinate in amount, if the nitrogen or proteid metabolism of the body is to be maintained at a level commensurate with true physiological requirements. But there comes the ever-recurring question, Are the lower proteid standards quite safe to follow? Are we warranted in turning aside from the teachings based on the habits and customs of mankind? Many reasons have already been presented which seemingly justify an affirmative answer, while the experimental results and the observations on various groups of people, covering years of time, speak with no uncertainty regarding the element of safety, and indicate clearly that the absolute proteid requirement of the body is quite small; much smaller indeed than the amount of proteid food consumed by the average individual would seemingly imply.
Probably the most striking evidence, certainly of an experimental nature, so far presented against the safety of a relatively low proteid diet for man is that based on the results of several studies made to ascertain the effect of a reduced proteid intake on so-called high proteid animals. Animal kind may be divided into three groups according to the nature of their food, viz., high proteid feeders, such as carnivorous animals in general, of which the dog is a good type; omnivorous or moderate proteid consumers, to which class man belongs; and low proteid consumers, such as herbivorous animals. Three series of experiments have been reported by independent workers on the effects of reducing the amount of proteid food in the diet of dogs. The results of these experiments were of such a character that it has come to be understood that animals of this type cannot exist for any great length of time on a low proteid diet. It is affirmed that in a relatively short period the animals reach such a state that232 they either die, or are in such poor condition that they must be fed a more liberal amount of proteid to maintain them alive. The explanation offered is that the low proteid diet results “in a loss of the power of absorption from the intestinal tract, caused apparently by a change in the condition of the epithelial cells, as well as by a diminished secretion of the digestive juices.”
The argument based on this evidence is that while a high proteid animal feels at once, or almost immediately, the deleterious effect of a reduction in the amount of proteid food, an omnivorous animal may be more tardy in manifesting the injurious action, which, however, is sure to follow sooner or later from any material reduction of proteid below the customary standards. In other words, man as a moderate proteid consumer can endure for a time even large reductions in the amount of proteid food, but eventually there will be manifested some of the disastrous results obtained with dogs. Here, we have a somewhat serious indictment, one that merits careful consideration. To be sure, it may be objected that between dog and man there is a wide gulf, and that there is no justification for assuming that these two types of animal life have anything in common. Still, the experience of many years has taught the physiologist that much light can be thrown upon the processes of higher types of life by a study of what occurs in lower forms, and on the subject of nutrition any one of experience would hesitate to cast out of court the evidence gathered from observation of what occurs among the higher animals. It will be the part of wisdom, therefore, to scrutinize somewhat carefully the character of this evidence obtained from a study of the behavior of dogs toward a low proteid diet.
The first series of experiments was made in 1891 by the late Immanuel Munk of Berlin, privat docent of physiology at the University, followed by further experiments in233 1893.67 Four dogs in all were studied. The diet made use of was “fleischmehl” (dried meat ground to a powder), fat (suet), and rice boiled together with water. We may refer briefly to the details of one experiment. The dog weighed 10.4 kilograms, and at first was given a daily diet composed of 85 grams of rice, 29 grams of fat, and 30 grams of the flesh meal. This ration contained 30.3 grams of proteid, 31 grams of fat, and 66 grams of carbohydrate, with a total fuel value of 663 calories, or 63 calories per kilogram of body-weight. On this diet, there was at the outset a slight loss of body-weight, after which both body equilibrium and nitrogen equilibrium were practically maintained. After this preliminary period of three weeks, the day’s diet was altered by replacing 15 grams of the proteid by 15 grams of rice, so that the daily ration consisted of 15.3 grams of proteid (with 2.42 grams of nitrogen), 31 grams of fat, and 81 grams of carbohydrate, with essentially the same fuel value per kilo of body-weight as before. Later, the fuel value of the food was further increased by raising the amount of rice to 125 grams per day, the day’s ration then consisting of 15.5 grams of proteid, 37 grams of fat, and 96 grams of carbohydrate, with a total fuel value of 780 physiological heat units, or 78 calories per kilo. On this diet, nitrogen equilibrium was maintained and the animal gained somewhat in body-weight. By the seventh week, however, Munk reports that the animal began to show signs of change; there was loss of appetite, absorption of the daily food was impaired, both proteid and fat failing in large degree to be utilized, while nitrogen equilibrium could no longer be maintained. This condition continued during the next week, aggravated by vomiting and accompanied by loss of strength and vigor. At the beginning of the tenth234 week of this low proteid ration, the animal was in a very poor condition, with complete loss of appetite, little inclination to take food, etc. On feeding a liberal diet of fresh meat, as much as 250 grams per day, with some fat (50 grams a day), the animal speedily recovered its appetite, and in a short time was in normal condition, absorption of food and utilization of the same being as complete as at the beginning of the experiment.
It is not necessary to give further details bearing on the three additional experiments. It will suffice to quote the general conclusions which Munk drew from the various results obtained, viz., that a low proteid intake in the case of dogs causes a loss of appetite, weakness, vomiting, etc., while body-weight and nitrogen equilibrium are difficult or impossible to maintain. More specifically, Munk’s observations led him to state that for dogs of ten kilograms body-weight a daily intake of 0.255 gram of nitrogen per kilo of body-weight is not sufficient to maintain the normal condition of the body, even when the fuel value of the day’s food amounts to more than 100 calories per kilo. In order to have the animal continue in nitrogen and body equilibrium, the daily food must contain at least 0.31 gram of nitrogen per kilogram of body-weight, with sufficient non-nitrogenous food to yield over 100 calories per kilo.
Let us now pass to the experiments made by Rosenheim,68 which were carried on at about the same date as Munk’s. In the first experiment, the dog weighed 11.3 kilograms, and was fed daily a low proteid ration having a fuel value of 1447 calories and containing 2.825 grams of nitrogen. This ration was reduced in a short time to a still lower plane, viz., to 1066235 calories and 2.525 grams of nitrogen daily. The food as then given was composed of 170 grams of rice, 50 grams of fat, and 25 grams of chopped meat, on which the dog gained weight and preserved nitrogen equilibrium. For six weeks, or thereabouts, the animal maintained its normal condition, after which it began to show symptoms of a general disturbance, with lack of appetite and weakness accompanied by a condition of icterus. Addition of meat extract to the diet to improve the flavor was without any appreciable effect. During the next two weeks, the condition of the animal steadily grew worse, although the body-weight remained practically stationary and nitrogen equilibrium was maintained. A week later, the animal died in a condition of exhaustion, without having manifested any symptoms of disturbed metabolism. There was found a marked catarrhal condition of the mucous membrane of the gastro-intestinal tract, with a fatty degeneration or metamorphosis of the glandular apparatus, but nothing sufficiently specific to account for the peculiar manner of death.
A second experiment with a dog of 5.8 kilograms, fed on meat, fat, and rice, led to essentially the same results as the preceding experiment. At the end of the first month, there appeared indications that the animal was not well, loss of appetite being marked, with disturbance of the stomach accompanied by occasional vomiting. These symptoms disappeared quickly when the animal was given for a few days large quantities of meat. On returning to the original low proteid diet, with its large content of rice, the symptoms gradually reappeared. At the end of two months the animal had again lost its appetite, and before the end of the fifth month the subject was dead. Post-mortem examination showed especially a strong fatty degeneration of the epithelial cells of the mucous membrane of the stomach and intestine. Rosenheim concludes that a diet poor in proteid is unhealthful for dogs, and that a daily ration containing even 0.32 gram of nitrogen236 per kilogram of body-weight, and with a fuel value of 110 calories per kilo, is not sufficient to maintain the animal in a condition of health.
The next series of experiments was made by Jägerroos69 of Finland. This investigator was evidently impressed by the unfavorable and monotonous character of the diet made use of by the preceding investigators, and sought to introduce a little variety, recognizing also that with a carnivorous animal it is difficult to reduce the proteid to a low level and maintain the necessary fuel value, without introducing foodstuffs to which the animal is wholly unaccustomed. In the first experiment, the dog had a body-weight of 5.77 kilograms, and at the beginning was fed daily 40 grams of meat and 100 grams of sugar, equal to 0.31 gram of nitrogen and 80 calories per kilo of body-weight. The experiment continued for eight months, sugar being replaced in part by butter, and occasionally bread, fat, and wheat meal being used in proper amount to yield the given nitrogen and fuel values. During the last five months, the intake of nitrogen per day averaged 0.29 gram per kilo, with a fuel value amounting to 89 calories daily per kilo of body-weight. During this period, the animal maintained a plus nitrogen balance for a large part of the time. The experiment was then continued for two months longer, with a gradual diminution in the nitrogen of the food and in the fuel value, the animal dying at the end of the tenth month.
In a second experiment, the dog made use of weighed at the beginning 11.97 kilograms. During the first five months, the average intake of nitrogen amounted daily to 0.29 gram per kilo, while the average fuel value of the food (meat, fat, and sugar) was 76 calories per kilo daily. In the middle of the seventh month the animal was quite ill, with poor appetite,237 vomiting, etc. Body-weight began to fall off, and the animal soon died. With both of these animals, the experiment ended suddenly by a sharp and short illness.
Jägerroos, however, believed that both animals died from a severe case of infection, and not as the result of the diminished intake of proteid. This view was fully substantiated, in his opinion, by the evidence furnished on bacteriological and morphological examination. There was no pathological alteration and no fatty degeneration in the intestinal epithelium; nothing to indicate any connection between the lowered proteid intake and the death of the animal. To be sure, the long-continued diet poor in nitrogen might have diminished the power of resistance of the body, but no proof of this is offered. There was indicated merely a simple infection, as shown by the presence of Streptococcus and Bacterium coli communis in the blood. But, as Jägerroos states, one might well conceive of a lowered power of resistance on the part of the body, due not to any change in diet, but to the long-continued confinement in a cage with the enforced inactivity and lack of freedom. It is to be noted, furthermore, that here there was no sign of a gradual and progressive weakening of the body, no indication of any disturbance of the digestive tract with diminished power of absorption of either fat or proteid. On the contrary, there was a sudden and sharp attack of some infectious disease by which the animals quickly succumbed. Jägerroos was of the opinion that in the absence of this infection the animals would have continued to live for a long period of time.
If a low proteid diet works so inimically on high proteid animals as Munk and Rosenheim thought, it would naturally be expected that the small proteid ration followed so long by Jägerroos would have resulted in the appearance of marked symptoms, at least a gradual and persistent falling off in body-weight, inability to maintain nitrogen equilibrium, etc.; but none of these things occurred. In Munk’s first experiment,238 the animal was given no fresh meat whatever during four weeks. Is it not quite possible that in the abrupt cutting off of this wonted form of food a disturbance may have been set up in the gastro-intestinal tract, which paved the way for the more serious results that followed? Jägerroos used only fresh, uncooked meat in his experiments, and laid great stress upon the importance of not departing any more than was necessary from the accustomed form of diet. The writer is strongly of the opinion that sufficient stress has not been laid upon this phase of the subject. A satisfactory diet for dog as for man must meet ordinary hygienic requirements; it must not only be sufficient in amount, but it must be easily digestible, of accustomed flavor, appealing to eye, nostrils, and palate, with reasonable variation occasionally and of moderate volume. With due regard to these conditions, I believe with Jägerroos that not much attention need be paid to the proportion of nitrogen therein, for however small the amount it will be found sufficient to meet the needs of the body.
These are the results, collectively, so frequently used to point a moral for man: Beware of the possible danger of reducing the consumption of proteid food below the commonly accepted dietary standards! We must admit, however, that there is a woeful lack of agreement in these results, and it is difficult to prevent a shadow of doubt from creeping over us as we try to depict for ourselves the way in which a low proteid ration exerts its deleterious effect on dogs. I do not believe that radical changes in diet, whether they involve increase or decrease in total quantities, or in specific elements of the diet, can be made suddenly without danger of some disturbance of the gastro-intestinal tract or other parts of the economy, either in dog or man. It is reasonable to believe also that a high proteid feeder, like a dog, with his more limited dietary, will be far more sensitive to great changes than omnivorous man with his wider range of foodstuffs. Moreover,239 there is just as good ground for believing that in any animal, excess of proteid is as dangerous as a low proteid diet. Too great a disturbance in the nutritive balance, whether it involves excess or reduction in the amount of a given foodstuff, is liable to be attended with serious disturbance in any sensitive organism.
In illustration of these statements, we have some recent results obtained by Watson and Hunter70 upon the influence of diet on growth and nutrition. These investigators find that young rats—two and a half months old—when fed upon a diet composed exclusively of horse-flesh, which is chiefly proteid matter with some fat, succumb very quickly, for some reason. Of fourteen young rats fed on this meat diet, six died on the third day. On the morning of this day, as the authors state, “the rats appeared to be in their usual health, but an hour after feeding one of them was lying on its side apparently unconscious. In a few minutes others were affected. They appeared to be paralyzed, they felt cold to the touch, exhibited symptoms of tetany, and speedily became unconscious. Six succumbed within half-an-hour. Of the remainder, some showed similar symptoms, although in less degree, and they recovered when the diet was changed to bread and skim milk.” After two days of the so-called normal diet, composed of bread and skim milk, the remaining eight rats were again placed on an exclusive meat diet. They appeared now to have acquired a certain degree of immunity, for although they exhibited symptoms of deranged nutrition, these were gradually recovered from and they gained in weight. At the end of the eighth month, five of the animals were still alive and in apparent good health, but their growth was permanently stunted. With an exclusive diet of ox-flesh, young rats were much240 more liable to thrive, although their growth was distinctly retarded.
This difference in the behavior of the animals towards the two forms of proteid food is to be attributed to the fact that ox-flesh contains more fat than horse-flesh, and consequently the diet with this form of meat was less exclusively proteid in character. Further, there were some indications that horse-flesh is less digestible than ox-flesh. Another fact, showing the far-reaching effect of a distinctly unphysiological diet, is the marked influence of pure meat food on the progeny. Thus, of 93 rats born of meat-fed parents only 19 were alive at the end of two months, while of 97 young born of bread and milk-fed rats, 82 were alive and in apparent health at the end of the same period.
As illustrating how foods that have, superficially at least, approximately the same chemical composition may react differently in the animal body we have the observations of Watson on rats fed with porridge, made by boiling oatmeal with water and skim milk, as contrasted with a diet of bread and skim milk, the two diets having essentially the same composition. Of fourteen young rats fed exclusively on porridge, all, with the exception of two that were withdrawn, succumbed within five months, while the bread and milk-fed animals thrived as usual. Adult rats, however, can live for prolonged periods and maintain their weight on a porridge diet. It is believed that the difference in the behavior of young rats to these two closely allied forms of diet, is due to a difference in the digestibility of the food, the porridge being presumably less readily digested by the young animals than bread. With the more fully developed digestive powers of the adult animals, however, this difference in availability practically disappears as a potent factor in their nutrition. Finally, mention may be made of the fact that a pure rice diet, notably deficient in proteid, arrests the growth of young rats and leads to a fatal241 issue within three months, while adult rats placed on such a diet lose weight rapidly and die in about the same time. All of these facts bearing on the nutrition of animals quite remote from man have significance as showing how any wide departure from a physiological diet, for that particular species or type, may lead to very undesirable results, and they warn us not to be too hasty in drawing far-reaching conclusions and sweeping deductions from a few experiments with a given species of animal.
Recurring now to the experiments made with dogs, there is certainly suggested an element of danger in a low proteid diet, which, if the experiments are taken at their face value and the conclusions derived therefrom applied to man, needs careful consideration. Jägerroos plainly was not inclined toward the belief that a low nitrogen intake was the cause of the unfortunate results that attended his experiments. Still, his animals did die from some cause, and thereby his position was weakened. Munk and Rosenheim, on the other hand, from their experiments were apparently convinced that a low proteid intake was inimical to dogs, and it will be remembered Rosenheim concluded that “a daily ration containing even 0.32 gram of nitrogen per kilogram of body-weight, and with a fuel value of 110 calories per kilo, is not sufficient to maintain the animal in a condition of health.” If this is really true, there is some ground for the arguments advanced by critical writers regarding the general subject of nitrogen requirements of man. The evidence and the arguments, however, have always seemed to the present writer frail and faulty; but recognizing the hold they have taken on physiologists and the way they are usually applied to man, I have attempted to test the matter experimentally under conditions which would yield trustworthy and conclusive results.
The question how far results obtained with dogs can be applied safely to man may be open to discussion, but we must242 first be sure of our facts before arguments or conclusions of any kind are warranted. It is to be remembered that dogs are as sensitive in many ways as man, and no physiological experiment covering a long period of time can be carried out with any hope of success unless there is due regard for proper hygienic conditions, some degree of variety in diet, and reasonable opportunities for fresh air and occasional exercise. I fancy that even the most vigorous and hardy man, if confined for six consecutive months in a room just large enough to furnish requisite air-space and to permit of extending his body at full length, would find himself at the end of such a period in a condition far from healthful, even though there were perfect freedom of choice in diet. If, however, there were added to the above conditions monotony in diet extending through many months, there would be no occasion for surprise if the individual lost appetite and strength, and showed signs of disturbance of the gastro-intestinal tract.
It is doubtful if there is full appreciation of the possible effect of monotony, in the ordinary dietary experiments on dogs. Man quickly feels the effect; the sportsman camping in the woods by brook or lake enjoys his first meal of speckled trout and has no thought of ever becoming tired of such a delicacy; but as trout cooked in various ways continue to be placed before him three times a day, and with perhaps very little else, he soon passes into a frame of mind where salt pork would be a luxury, and where he would prefer to go hungry rather than eat the delicacy, if indeed he has appetite to eat anything. Is it strange that dogs confined in cages barely large enough to permit of their turning around, and fed day after day and month after month with exactly the same amount of desiccated meat, fat, and rice, should show signs and symptoms, if nothing worse, of disturbed nutrition? It is necessary in experiments of this kind that the animals be confined for given periods, at least, since otherwise it243 would be impossible to determine the extent of nitrogen excretion and the rate of proteid katabolism, etc. It is possible, however, to limit the time of close confinement to, say, ten consecutive days, this to be followed by a like period of comparative freedom, thus insuring opportunities for an abundance of fresh air and exercise.
The experiments of which I wish to speak, and which had for their object a study of the effect of low proteid diet on dogs, as types of high proteid animals, were carried out at our laboratory in the Sheffield Scientific School and were made possible by liberal grants from the Carnegie Institution of Washington, thus providing means for securing the requisite number of chemical assistants. The experiments were conducted on a somewhat large scale, over twenty dogs being made use of, while many of the experiments extended through a full year. The results in their entirety are not yet ready for publication, but I am able to present in a general way observations on six dogs, which will serve as an ample illustration of what may be expected with high proteid animals when living on a low proteid diet under healthful conditions. All of the six dogs whose cases are here presented were fed on a mixed diet, with some fresh meat each day; bread, cracker dust, milk, lard, and rice being the other foods drawn upon to complete the dietary. The animals were fed twice a day, each meal being accurately weighed and of definite chemical composition. A large, light, and airy room, kept scrupulously clean, and in the winter time properly heated by steam, served as their main abiding place. In this room were a suitable number of smaller compartments, the walls of which were composed of open lattice work (of iron), so as not to interfere with light or air, and yet adequate to keep the dogs apart. These compartments were not cages in the ordinary sense, but were truly large and roomy. The entire floor under the dogs was composed of metal, the joints all soldered,244 the floor being sloped to a metal gutter in front so that all the compartments could be flushed out each morning and kept sweet and clean. In pleasant weather, immediately after their first meal, the dogs were taken out of doors to a large enclosure near by, where they were allowed perfect freedom until about four o’clock, when they were taken in for their second meal (between four and five o’clock in the afternoon). The outdoor enclosure was inaccessible to every one except the holder of the key, and the dogs while there were wholly free from annoyance. Once every month, during a period of ten consecutive days, each dog was confined in the metabolism cage so as to admit of the collection of all excreta, in order to make a determination of the nitrogen balance. Practically, therefore, each dog was in close confinement only one-third of the month, the remaining two-thirds being spent in much more congenial surroundings. I have entered thus fully into a description of the conditions prevailing, because I deem them exceedingly important, and because therein undoubtedly lies the explanation of the striking contrast between our results and those of the earlier investigators of this subject.
In considering the outcome of our experiments, it may be wise to enter into some detail concerning the first case to be presented. The animal employed in this experiment was designated as No. 5, and weighed on July 27, 1905, 17.2 kilograms; it was apparently full grown, but was thin and had the appearance of being underfed. At first, it was given daily 172 grams of meat, 124 grams of cracker dust, and 72 grams of lard, the day’s ration containing 8.66 grams of nitrogen and having a fuel value of 1389 calories.71 These figures are equivalent to 80 calories, and 0.50 gram of nitrogen, per kilogram of body-weight. The animal took kindly to the diet,245 but on August 3 it refused to eat and seemed to have a little fever. The next day it was better, but for the three following days its appetite was poor, and only a portion of the daily food was eaten. Body-weight began to fall off, and was soon at 15.5 kilograms. On the 7th of August, a dose of vermifuge was given, after which the appetite returned and the animal appeared in good spirits. From this time forward it seemed in perfect health, with good appetite, and showed the usual vivacity and playfulness of dog-kind. The diet as specified was continued unchanged until August 25, a balance experiment covering a period of ten days, from the 15th to246 the 24th of August inclusive, being carried out, in which the nitrogen of the intake was compared with the output for each day. From the accompanying table, where are given the average values of all the balance periods of the experiment, it is to be seen that during this first period the animal was laying on or gaining an average of 2 grams of nitrogen per day.
SUBJECT No. 5. DAILY AVERAGES
Date. |
Body- weight. |
Food. |
Output. |
Nitro- gen Balance + or – |
||||
|---|---|---|---|---|---|---|---|---|
| Total Nitro- gen. |
Nitro- gen per Kilo Body- weight. |
Fuel Value per Kilo Body- weight. |
Nitro- gen through Excre- ment. |
Nitro- gen through Hair. |
||||
1905 |
kilos |
grams |
gram |
calories |
grams |
gram |
gram |
grams |
| Aug. 15–Aug. 24 | 15.8 |
8.66 |
0.54 |
87.3 |
5.44 |
0.70 |
0.52 |
+2.00 |
| Sept. 6–Sept. 15 | 17.1 |
4.76 |
0.27 |
72.4 |
3.41 |
0.32 |
0.48 |
+0.55 |
| Oct. 8–Oct. 17 | 17.6 |
4.76 |
0.27 |
71.8 |
3.54 |
0.54 |
0.49 |
+0.19 |
| Nov. 22–Dec. 1 | 16.9 |
4.77 |
0.28 |
72.0 |
3.76 |
0.39 |
0.32 |
+0.30 |
1906 |
||||||||
| Jan. 2–Jan. 11 | 17.2 |
4.07 |
0.23 |
72.0 |
3.19 |
0.54 |
0.35 |
–0.01 |
| Jan. 30–Feb. 8 | 18.0 |
4.07 |
0.23 |
69.0 |
2.87 |
0.54 |
0.62 |
+0.04 |
| Feb. 27–Mar. 8 | 18.2 |
5.18 |
0.28 |
73.0 |
3.69 |
0.66 |
0.74 |
+0.09 |
| Mar. 27–Apr. 5 | 18.3 |
5.23 |
0.28 |
73.0 |
3.66 |
0.84 |
0.48 |
+0.25 |
| Apr. 24–May 3 | 19.1 |
5.22 |
0.27 |
68.0 |
3.76 |
0.38 |
0.48 |
+0.60 |
| May 22–May 31 | 19.4 |
5.22 |
0.26 |
65.0 |
3.44 |
0.31 |
0.48 |
+0.99 |
| June 17–June 26 | 20.0 |
5.24 |
0.26 |
67.0 |
3.50 |
0.71 |
0.48 |
+0.55 |
On August 25, a radical change was made in the diet, by reducing the amount of meat to 70 grams daily, thereby lowering the intake of nitrogen to 4.76 grams, or 0.27 gram per kilo of body-weight; the cracker dust and lard being kept at essentially the same levels as before. This diet was continued through the next balance period, the dog in the meantime gaining in body-weight, and showing for the second balance period an average gain by the body of half a gram of nitrogen per day. The food was then altered by substituting bread for the cracker dust, but so adjusted that the nitrogen and fuel values of the day’s food remained practically unchanged. There was still, however, a gain in body-weight and a slight gain in body nitrogen. At the close of the third balance period, the diet was again altered, one-half of the meat being replaced by milk, while cracker dust was substituted for the bread. The morning meal consisted of 170 grams of milk, 86 grams of cracker dust, and 18 grams of lard, while the afternoon meal was composed of 35 grams of meat, 63 grams of cracker, and 35 grams of lard. The day’s ration, however, still contained 4.76 grams of nitrogen and had a fuel value of 1249 calories. This diet was maintained until November 20, when the animal was again placed on a daily ration of meat (69 grams), bread (166 grams), and lard (80 grams), with a total fuel value of 1228 calories and 4.77 grams of nitrogen. This was continued until December 2, the dog still showing a plus nitrogen balance, but with a little loss in body-weight. On December 2, the diet was again247 changed by substituting milk for a portion of the meat, but the nitrogen and fuel values were maintained at the same level as before. After a week, December 9, the food was modified as follows: the morning meal contained 170 grams of milk, 110 grams of rice, and 11 grams of lard, while the afternoon meal was composed of 35 grams of meat, 81 grams of rice, and 30 grams of lard. The total nitrogen content of the day’s ration was 4.07 grams, while the fuel value was 1255 calories. At this time, the animal weighed 17.1 kilograms, consequently the intake of nitrogen had been reduced to 0.23 gram per kilo of body-weight, while the fuel value stood at 73 calories per kilogram. This diet was continued until February 9, the balance period, between January 2 and 11, showing that the animal was in nitrogen equilibrium, in spite of the material reduction in the intake of proteid, and that body-weight was increasing. The next balance period, January 30 to February 8, showed still further gain in weight with continuance of nitrogen equilibrium. On February 9, the diet was changed by returning to 70 grams of meat, 158 grams of cracker dust, and 60 grams of lard, with a daily intake of 0.28 gram of nitrogen per kilo of body-weight.
In this manner, the experiment was continued with frequent changes in the character of the diet, but always maintaining essentially the same values in nitrogen and calories as shown in the table, until June 27; having extended through just eleven months, with the animal at the close of the experiment still gaining in body-weight, with a steady plus balance of nitrogen, and with every indication of good health and strength. For ten months the animal lived with perfect comfort and in good condition on an average daily intake of 0.26 gram of nitrogen per kilogram of body-weight, and with an average fuel value of 70.3 calories per kilo. Further, it is to be observed that at no time during the ten months did the daily intake of nitrogen rise above 0.28 gram per kilo, while during one248 month it fell to 0.23 gram per kilo. Similarly, the fuel value of the daily food never exceeded 73 calories per kilo, while at times it dropped as low as 67 and 65 calories per kilo. That this diet was more than sufficient, both in nitrogen and fuel value, is indicated by the steady increase in body-weight and by the plus nitrogen balances observed in most of the periods throughout the experiment. Indeed, with the comparatively low degree of muscular activity which this animal was accustomed to, it would have been unwise to have kept the subject much longer on a diet so rich as the above, since there would have been danger of detriment to its health and good condition. When these results are contrasted with the statements of Munk and Rosenheim, the latter of whom found that even 0.32 gram of nitrogen and 110 calories per kilo were insufficient to maintain dogs in a condition of health, it is plain that for some reason our results are quite at variance with their findings.
The accompanying photographs, taken on August 19, 1905, February 27, April 24, and at the close of the experiment on June 27, 1906, show the appearance of the animal at the respective dates, and indicate more clearly than words can express the actual condition of the animal.
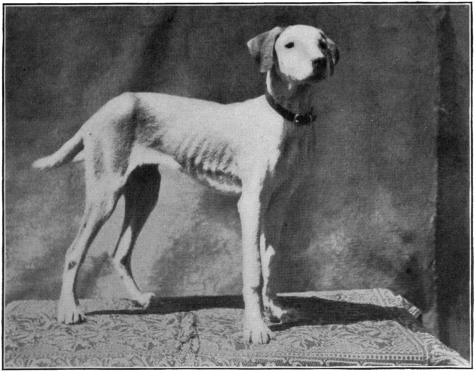
Subject No. 5. August 19, 1905

Subject No. 5. November 18, 1905

Subject No. 5. April 24, 1906

Subject No. 5. June 27, 1906
Turning now to a second subject, designated as dog No. 3, the experiment with which lasted for nearly an entire year, the following general statements may be made. The animal was a small black and white fox terrier, weighing on July 6, 1905, 6.5 kilograms. It was a nervous, affectionate little creature, far less phlegmatic than the animal just described, always on the alert for a petting, and unceasingly active. For these reasons, it seemingly required per kilogram of body-weight a little more food than the preceding animal; a fact also in harmony with the general law that small animals, per unit of body-weight, need more food than larger ones. The diet made use of was of the same general character as employed249 with the preceding animal, and was changed from time to time to give requisite variety and to insure freedom from too great monotony. The accompanying table, showing daily averages during the twelve balance periods, gives all necessary information regarding the outcome of the experiment.
SUBJECT No. 3. DAILY AVERAGES
| Date. | Body- weight. |
Food. | Output. | Nitro- gen Balance + or – |
||||
|---|---|---|---|---|---|---|---|---|
Total Nitro- gen. |
Nitro- gen per Kilo Body- weight. |
Fuel Value per Kilo Body- weight. |
Nitro- gen through Kidneys. |
Nitro- gen through Excre- ment. |
Nitro- gen through Hair. |
|||
1905 |
kilos |
grams |
gram |
calories |
grams |
gram |
gram |
gram |
| July 18-July 28 | 6.8 |
5.88 |
0.84 |
79.0 |
5.58 |
0.43 |
0.05 |
–0.18 |
| Aug. 15-Aug. 24 | 7.1 |
3.44 |
0.49 |
77.4 |
3.35 |
0.17 |
0.13 |
–0.21 |
| Sept. 6-Sept. 15 | 6.9 |
2.11 |
0.30 |
80.0 |
1.93 |
0.21 |
0.07 |
–0.10 |
| Oct. 8-Oct. 17 | 6.9 |
2.10 |
0.30 |
80.0 |
1.83 |
0.20 |
0.07 |
0 |
| Nov. 22-Dec. 1 | 6.0 |
1.83 |
0.31 |
80.0 |
1.48 |
0.21 |
0.11 |
+0.03 |
1906 |
||||||||
| Jan. 2-Jan. 11 | 5.6 |
1.63 |
0.29 |
81.0 |
1.54 |
0.17 |
0.08 |
–0.16 |
| Jan. 30-Feb. 8 | 5.5 |
1.63 |
0.30 |
82.0 |
1.60 |
0.15 |
0.05 |
–0.17 |
| Feb. 27-Mar. 8 | 5.5 |
1.78 |
0.32 |
84.0 |
1.66 |
0.17 |
0.05 |
–0.10 |
| Mar. 27-Apr. 5 | 5.7 |
1.98 |
0.34 |
81.0 |
1.75 |
0.21 |
0.06 |
–0.04 |
| Apr. 24-May 3 | 5.7 |
1.98 |
0.34 |
83.0 |
1.68 |
0.13 |
0.13 |
+0.04 |
| May 22-May 31 | 5.8 |
1.98 |
0.34 |
80.0 |
1.77 |
0.13 |
0.11 |
–0.03 |
| June 17-June 26 | 6.0 |
1.98 |
0.33 |
77.0 |
1.53 |
0.21 |
0.07 |
+0.17 |
It will be observed that during the first three months the animal showed a tendency to gain in weight slightly, recalling that its initial weight on July 6 was 6.5 kilograms. Later, the weight fell off a little, but in March it showed an upward movement, though very gradual. With the amount of proteid food given, it is evident that the animal needed about 80 calories per kilo to maintain a condition of body-equilibrium.250 Nitrogen equilibrium was practically maintained throughout the larger portion of the twelve months, but evidently the animal required 0.31–0.33 gram of nitrogen per kilogram of body-weight. Attention may be directed, in view of the results reported by Munk regarding loss of the power of absorption and utilization of proteid food, to the figures showing the average daily output of nitrogen through the excrement. It is plain from the data presented, that this animal was not suffering from any trouble of this order; indeed, the utilization of proteid food throughout the entire experiment was exceedingly complete, as shown by the relatively small loss of nitrogen through the excrement, thus implying vigorous and unimpaired digestion, together with thorough absorption of the products formed.
The accompanying photographs show the appearance of the 251 animal on August 19, 1905, November 18, 1905, April 3 and June 27, 1906, the close of the experiment.

Subject No. 3. August 19, 1905

Subject No. 3. November 18, 1905

Subject No. 3. April 24, 1906

Subject No. 3. June 27, 1906
Passing now to the third subject, we have an experiment of somewhat shorter duration, viz., of nine months, but sufficiently long to afford ample opportunity for any deleterious effect to manifest itself. The initial weight of the dog, No. 13, was 14.5 kilograms on September 14. The lowest intake of nitrogen was 0.26 gram per kilo of body-weight per day, while the fuel value of the daily food was during one period reduced to 55 calories per kilo. A daily proteid consumption equalling 0.30 gram of nitrogen per kilo, with a total fuel value in the day’s food of 66–70 calories per kilo, was clearly quite sufficient to maintain nitrogen equilibrium and body-weight; indeed, toward the end of the experiment, the animal commenced to gain in weight quite noticeably on the above diet, and was laying by fairly large amounts of nitrogen252 daily. The accompanying table gives the average daily nitrogen exchange, etc., of the nine balance periods, while the photographs, taken on the dates indicated under each, show the appearance of the animal at various times.
SUBJECT No. 13. DAILY AVERAGES
| Date. | Body- weight. |
Food. | Output. | Nitro- gen Balance + or – |
||||
|---|---|---|---|---|---|---|---|---|
Total Nitro- gen. |
Nitro- gen per Kilo Body- weight. |
Fuel Value per Kilo Body- weight. |
Nitro- gen through Kid- neys. |
Nitro- gen through Excre- ment. |
Nitro- gen through Hair. |
|||
1905 |
kilos |
grams |
gram |
calories |
grams |
gram |
gram |
gram |
| Sept. 24-Oct. 3 | 14.0 |
7.22 |
0.52 |
86.0 |
6.40 |
0.71 |
0.19 |
–0.08 |
| Nov. 5-Nov. 14 | 13.0 |
4.78 |
0.35 |
80.0 |
4.29 |
0.37 |
0.25 |
–0.13 |
| Dec. 19-Dec. 28 | 13.4 |
3.70 |
0.27 |
72.0 |
2.86 |
0.49 |
0.13 |
+0.22 |
1906 |
||||||||
| Jan. 16-Jan. 25 | 14.1 |
3.72 |
0.26 |
70.0 |
3.16 |
0.61 |
0.16 |
–0.21 |
| Feb. 13-Feb. 22 | 14.3 |
4.26 |
0.30 |
78.0 |
3.54 |
0.67 |
0.37 |
–0.32 |
| Mar. 13-Mar. 22 | 14.1 |
3.62 |
0.26 |
55.0 |
3.29 |
0.46 |
0.14 |
–0.27 |
| Apr. 10-Apr. 19 | 14.2 |
4.59 |
0.32 |
73.0 |
2.84 |
0.51 |
0.10 |
+1.14 |
| May 8-May 17 | 14.2 |
4.59 |
0.32 |
71.0 |
3.56 |
0.48 |
0.18 |
+0.37 |
| June 5-June 14 | 15.3 |
4.58 |
0.30 |
66.0 |
2.98 |
0.55 |
0.28 |
+0.77 |

Subject No. 13. January 2, 1906

Subject No. 13. February 27, 1906

Subject No. 13. April 24, 1906

Subject No. 13. June 19, 1906
Results of the same general tenor with dogs No. 15 and No. 20 are seen in the appended tables, while the accompanying photographs testify clearly to the general good condition of the animals up to the end of the experiments. In No. 20 particularly, the great gain in body-weight is to be noted, even though the fuel value of the food was reduced as low as 64 calories per kilo, with the nitrogen intake at 0.28 gram per kilo daily. Plainly, the day’s food could have been diminished still more, with perfect safety to both body and nitrogen equilibrium.
SUBJECT No. 15. DAILY AVERAGES
Date. |
Body- weight. |
Food. |
Output. |
Nitro- gen Balance + or – |
||||
|---|---|---|---|---|---|---|---|---|
Total Nitro- gen. |
Nitro- gen per Kilo Body- weight. |
Fuel Value per Kilo Body- weight. |
Nitro- gen through Kid- neys. |
Nitro- gen through Excre- ment. |
Nitro- gen through Hair. |
|||
1905 |
kilos |
grams |
gram |
calories |
grams |
gram |
gram |
gram |
| Nov. 5-Nov. 14 | 9.2 |
3.35 |
0.36 |
82.0 |
2.95 |
0.11 |
0.14 |
+0.15 |
| Dec. 19-Dec. 28 | 8.9 |
2.61 |
0.30 |
75.0 |
2.47 |
0.12 |
0.12 |
–0.10 |
1906 |
||||||||
| Jan. 16-Jan. 25 | 8.7 |
2.60 |
0.30 |
79.9 |
2.15 |
0.21 |
0.16 |
+0.08 |
| Feb. 13-Feb. 16 | 8.5 |
2.61 |
0.30 |
82.0 |
2.37 |
0.20 |
0.15 |
–0.11 |
| Mar. 13-Mar. 22 | 8.7 |
2.82 |
0.32 |
80.0 |
2.68 |
0.17 |
0.19 |
–0.22 |
| Apr. 10-Apr. 19 | 9.0 |
2.80 |
0.31 |
82.0 |
2.14 |
0.26 |
0.09 |
+0.31 |
| May 8-May 17 | 9.5 |
2.83 |
0.30 |
75.0 |
2.26 |
0.30 |
0.12 |
+0.15 |
| June 5-June 14 | 10.2 |
2.81 |
0.27 |
70.0 |
2.26 |
0.28 |
0.24 |
+0.03 |

Subject No. 15. January 2, 1906

Subject No. 15. February 27, 1906
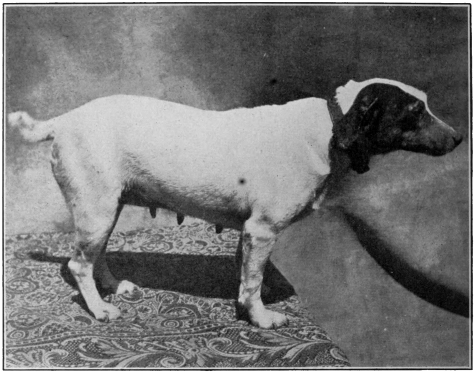
Subject No. 15. April 24, 1906

Subject No. 15. June 19, 1906
SUBJECT No. 20. DAILY AVERAGES
Date. |
Body- weight. |
Food. |
Output. |
Nitro- gen Balance + or – |
||||
|---|---|---|---|---|---|---|---|---|
Total Nitro- gen. |
Nitro- gen per Kilo Body- weight. |
Fuel Value per Kilo Body- weight. |
Nitro- gen through Kid- neys. |
Nitro- gen through Excre- ment. |
Nitro- gen through Hair. |
|||
1905 |
kilos |
grams |
gram |
calories |
grams |
gram |
gram |
gram |
| Dec. 6-Dec. 15 | 15.9 |
8.35 |
0.52 |
82.0 |
6.03 |
0.74 |
0.38 |
+1.20 |
1906 |
||||||||
| Jan. 16-Jan. 25 | 16.4 |
4.47 |
0.27 |
73.0 |
3.61 |
0.55 |
0.15 |
+0.16 |
| Feb. 13-Feb. 22 | 17.2 |
4.45 |
0.25 |
72.0 |
3.92 |
0.36 |
0.13 |
+0.04 |
| Mar. 13-Mar. 22 | 17.4 |
5.00 |
0.28 |
72.0 |
5.49 |
0.33 |
0.10 |
–0.92 |
| Apr. 10-Apr. 19 | 18.4 |
5.60 |
0.30 |
69.0 |
4.88 |
0.52 |
0.18 |
+0.02 |
| May 8-May 17 | 19.6 |
5.58 |
0.28 |
69.0 |
3.85 |
0.75 |
0.38 |
+0.60 |
| June 5-June 14 | 19.7 |
5.59 |
0.28 |
64.0 |
4.69 |
0.45 |
0.40 |
+0.05 |
253

Subject No. 20. January 2, 1906

Subject No. 20. February 27, 1906
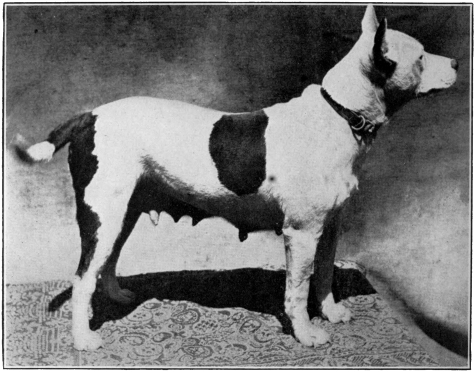
Subject No. 20. April 24, 1906

Subject No. 20. June 19, 1906
The illustrations so far presented, with the general agreement in the character of the results, might perhaps be interpreted as indicating that there is no difficulty whatever in bringing a high proteid consumer, like a dog, down to a low level of proteid consumption. This, however, would be a false impression. Much depends upon the character of the proteid food, at least where any attempt at rapid change is made, for a certain modicum of meat or other animal food seems a necessary part of the daily diet if health and strength are to be maintained. A dog transferred suddenly from a daily ration in which meat and milk are conspicuous elements to a diet in which these are wholly wanting is very liable to show disturbing symptoms almost immediately. One case may be cited in illustration of these statements. On September 29, 1905, dog No. 17, weighing 18.2 kilos, was placed on a daily diet composed of 70 grams of fresh meat, 442 grams of milk, 300 grams of bread, and 28 grams of lard. This ration contained 9.06 grams of nitrogen and had a fuel value of 1465 calories, or 0.5 gram of nitrogen and 80 calories per kilogram of body-weight. On October 11, the animal weighed 18.6 kilograms and was in perfect condition. On the 13th, the meat was reduced to 34 grams per day, but the milk was increased in amount so as to maintain the same nitrogen intake and fuel value as before. This diet was continued until November 3, a balance experiment covering ten days from October 22 to the 31 inclusive, showing that the animal was laying by a little nitrogen. On November 3, the diet was changed to milk, bread, and lard, the fuel value being maintained at 80 calories per kilo daily, while the nitrogen intake was reduced to 0.30 gram per kilo. On this diet, the animal seemed to thrive perfectly, and at the end of two weeks showed a body-weight of 18.2 kilograms. November 19, the milk was withdrawn, the bread being increased so as to keep the daily nitrogen intake and the fuel value unchanged. The day’s food was now composed of bread and lard solely, but, as just stated, the nitrogen and fuel values were unaltered. In four days’ time, however, a change began to creep over the254 animal; the appetite diminished, and there was apparent a condition of lassitude and general weakness which deterred the animal from moving about as usual.
During the next week the animal grew steadily worse, and would eat only when coaxed with a little milk or with bread softened with milk, the diet of bread and lard being invariably refused. There was marked disturbance of the gastro-intestinal tract; bloody discharges were frequent; the mucous membrane of the mouth was greatly inflamed and very sore; body-weight fell off, and the animal was in a very enfeebled condition. This continued until December 4, with every indication that the animal would not long survive, but by feeding carefully with a little milk and occasionally some meat, improvement finally manifested itself, and by December 18 there was good appetite, provided bread was not conspicuous in the food. Body-weight, which had fallen to 15.5 kilos, was being slowly regained, and on December 30 the animal was again placed on a weighed diet, consisting of 70 grams of meat, 442 grams of milk, 210 grams of cracker dust, and 10 grams of lard. This diet contained 8.26 grams of nitrogen and had a fuel value of 1330 calories, equivalent to 0.5 gram nitrogen and 80 calories per kilogram of body-weight. On January 12, 1906, the weight of the animal was 16.7 kilos, while in general condition there was nothing to be desired. The food was then modified by diminishing the amounts of meat and milk fed daily by one-half, thus reducing the nitrogen intake to 0.35 gram per kilo of body-weight, but maintaining the fuel value of the food at 80 calories per kilo. Under this régime, body-weight still increased, and on January 27 was 17.5 kilograms. A balance period, shown in the accompanying table, extending from January 30 to February 8, affords ample evidence that the body was laying by nitrogen.
255
SUBJECT No. 17. DAILY AVERAGES
| Date. | Body- weight. |
Food. |
Output. |
Nitro- gen Balance + or – |
||||
|---|---|---|---|---|---|---|---|---|
| Total Nitro- gen. |
Nitro- gen per Kilo Body- weight. |
Fuel Value per Kilo Body- weight. |
Nitro- gen through Kid- neys. |
Nitro- gen through Excre- ment. |
Nitro- gen through Hair. |
|||
1905 |
kilos |
grams |
gram |
calories |
grams |
gram |
gram |
gram |
| Oct. 22-Oct. 31 | 18.3 |
9.06 |
0.49 |
80.0 |
7.73 |
0.66 |
0.28 |
+0.39 |
1906 |
||||||||
| Jan. 30-Feb. 8 | 17.6 |
5.77 |
0.33 |
78.0 |
4.12 |
0.44 |
0.21 |
+1.00 |
| Feb. 27-Mar. 8 | 17.9 |
5.31 |
0.30 |
72.0 |
4.59 |
0.59 |
0.37 |
–0.24 |
| Mar. 27-Apr. 5 | 18.1 |
5.33 |
0.29 |
70.0 |
5.63 |
0.89 |
0.27 |
–1.52 |
| Apr. 24-May 3 | 18.4 |
5.90 |
0.32 |
68.0 |
5.06 |
0.49 |
0.30 |
+0.05 |
| May 22-May 31 | 18.6 |
5.90 |
0.31 |
67.0 |
5.25 |
0.53 |
0.43 |
–0.31 |
| June 17-June 26 | 19.9 |
5.89 |
0.29 |
70.0 |
4.29 |
0.39 |
0.28 |
+0.93 |
In all of the subsequent months, a small amount of meat was a part of the daily food, but as is seen from the table of balance periods, the total nitrogen intake and the fuel value of the food were reduced to even lower levels per kilogram of body-weight. Yet the animal gained steadily, until at the latter part of June the weight was considerably above that noted at the commencement of the experiment in the preceding October. Further, the animal was in nitrogen equilibrium or even gaining nitrogen, and in perfect condition of health and vigor, as is indicated by the accompanying photographs taken at the different periods stated. Especially to be emphasized is the fact that during the last six months of the experiment, the daily intake of nitrogen and the fuel value of the food were as low or even lower than in November, when the daily diet was limited to bread and lard. The disastrous result which showed itself at once on this latter diet, with all animal food excluded, was not due to low proteid or to deficiency in fuel value, but simply to the fact that the animal for256 some reason could not adjust itself to a simple dietary of bread and fat, although there was ample available nitrogen and fuel value for the body’s needs. Something was lacking, which meat or milk could supply, and this something was indispensable for the maintenance of the normal nutritional rhythm.

Subject No. 17. January 2, 1906

Subject No. 17. February 27, 1906

Subject No. 17. April 24, 1906

Subject No. 17. June 27, 1906
This is by no means an exceptional case, but we can cite many other examples of like results where the animal when restricted to a purely vegetable diet, such as bread, pea-soup, bean soup, etc., reinforced by an animal fat, quickly passed from a condition of health into a state of utter wretchedness, with serious gastro-intestinal disturbance. The results are not to be attributed to the lower utilization of the vegetable food, for the disastrous effect is too quickly manifest, and further, often shows itself when the animal plainly has a large store of available nutriment in its own tissues.
This experiment with dog No. 17 has been dwelt upon at some length, because it illustrates a very important principle in the nutrition of a high proteid and carnivorous animal. As before stated, it is not a question of high or low proteid simply, but involves possibly the more subtle question of the relative value of specific forms of proteid food. It will be noted that this statement is made somewhat guardedly, in harmony with the caution necessarily called for in view of our lack of knowledge regarding the possible need of the animal’s body for extraneous principles which only meat, milk, or other animal products can supply. Inorganic salts, nitrogenous extractives, and other substances without any appreciable fuel value, are quite likely to be of primary importance in controlling and regulating the various processes of the body, which combine to maintain the condition of normal nutrition. With a diet restricted to one or two vegetable products, it is quite conceivable that something may be lacking which the system demands, though it cannot be measured in terms of nitrogen or calories. It may be said that man thrives on a purely vegetable diet, but while this is unquestionably true, it must be remembered that man with his free choice of food has recourse, as a rule, to a large variety of vegetable products from many sources, and consequently there is great likelihood of his absorbing from these varied products such supplementary matters as may be needed. On this question, we are in a realm of doubt and uncertainty, but the possibilities suggested must not be ignored, for they may contain a germ of truth of the utmost importance. The fact remains, however, that a dog when restricted to a purely vegetable dietary does not thrive; a little animal food seems necessary to keep up health and strength, and this suffices even though the daily nitrogen intake and fuel value of the food are restricted to a level below that of the vegetable dietary.
With these facts before us, it is difficult to avoid the conclusion that some significance may attach to the specific nature of the proteid. Of course, we must not overlook the radical difference in dietary habits of man and dog. Man as an omnivorous creature has for generations been accustomed to partake largely of vegetable foods, and as a result his digestive tract and his system as a whole has become acclimated, as it were, to the nutritive effects of vegetable matter. Dogs, on the other hand, are typical carnivores, and their habits for generations have led in an opposite direction, so that their gastro-intestinal tracts and their systems have become accustomed to the effects of a diet in which animal food largely predominates. Whether these deeply ingrained characteristics are responsible in any large measure for the difference in behavior of man, on a purely vegetable diet, and dogs is open to question. It would certainly not be strange if such were the case, but as we look at the facts collected in our study of this subject, it is somewhat impressive to note how well dogs thrive on a relatively large amount of vegetable food, provided258 there is a modicum of animal food added thereto. In other words, these high proteid consumers are apparently quite able to utilize the vegetable foods, but there is something lacking in such a dietary which the body has great need of. Is it not quite possible, as already suggested, that the specific nature of the proteid counts for something in nutrition? The question cannot be answered definitely at present, but there are certain facts slowly accumulating which make the question a pertinent one in this connection.
Thus, it is becoming evident, as was pointed out in an earlier chapter, that the many proteid substances occurring in the animal and vegetable kingdoms are more or less unlike each other in their chemical make-up. They yield different decomposition products, or the same products in widely different proportion, when broken down by the action of hydrolyzing agents; and when we recall that the digestive enzymes of the body convert the proteids of the food into these same end-products, it is plain that in the assimilation and utilization of the proteid foodstuffs the body has to deal with these various chemical units. Hence, an animal suddenly restricted to a dietary in which all of the proteid is furnished by bread might be seriously incommoded, either by the excess of certain amino-acids resulting therefrom, or by a lack of certain other end-products to which its body is accustomed. As an example, we may take the three typical proteids of the wheat kernel, gliadin, glutenin, and leucosin, and note the very striking difference in the proportion of certain of the decomposition products of each, as reported by Osborne and Clapp.73
259
| Gliadin. | Glutenin. | Leucosin. | |
|---|---|---|---|
per cent |
per cent |
per cent |
|
| Leucin | 5.61 |
5.95 |
11.34 |
| Lysin | 0 |
1.92 |
2.75 |
| Arginin | 3.16 |
4.72 |
5.94 |
| Glutaminic acid | 37.33 |
23.42 |
6.73 |
| Ammonia | 5.11 |
4.01 |
1.41 |
| Aspartic acid | 0.58 |
0.91 |
3.35 |
| Tyrosin | 1.20 |
4.25 |
3.34 |
It is obvious from these figures that the three proteids of the wheat kernel are radically different from each other. Contrast, for example, the content of glutaminic acid in gliadin with the amount in leucosin. With such striking differences in chemical make-up, it is reasonable to assume that corresponding differences in physiological action or food values may exist. Further, “in respect to the amount of these amino-acids, leucosin more nearly resembles the animal proteins than the seed proteins thus far examined, and in this connection it is interesting to note that leucosin occurs chiefly if not wholly in the embryo of this seed and is probably one of its ‘tissue’ proteins, in contrast to the ‘reserve’ proteins of the endosperm of which gliadin and glutenin form the chief part” (Osborne and Clapp). In other words, animal proteids, such as those of meat, are characterized like leucosin by a small content of glutaminic acid and ammonia; while leucin, lysin, aspartic acid, and arginin are relatively more abundant. Until we know more on this subject, however, any broad generalization would be out of place, but certainly there is justification for the supposition that in these differences in chemical constitution are to be found explanation of some of the peculiarities common to certain varieties of proteid food. Wheat flour,260 aside from its starch, is composed mainly of glutenin and gliadin with their large content of glutaminic acid. Meat proteids, on the other hand, like leucosin, contain only a small fraction of this acid, and, with the other differences indicated, meat proteid and wheat proteid as food for dogs or other high proteid consumers may reasonably be expected to have at the least very unequal values. And if we go a step beyond this and suppose that in the formation of true tissue proteid or the living protoplasm of the cell, certain of these end-products of proteid decomposition are absolutely indispensable, we can easily picture for ourselves a dearth of such building stones in the long-continued use of a diet which lacks that particular proteid from which the necessary building stones can be split off in adequate number.
It has been said, notably by Munk, that in dogs fed for some time on a low proteid diet there is a diminished power of absorption from the intestinal tract, associated with weakened digestion. If it is true that a lowered proteid intake results in a diminished utilization of the ingested food, that efficiency in the digestion and absorption of foodstuffs is impaired, it can only be interpreted as meaning that some injurious influence has been exerted on the epithelial cells of the intestine or the adjacent gland cells. We have, however, failed to find any evidence of deleterious action in the dogs that we have experimented with, where due regard was paid to maintaining a diet suitable for the physiological needs of the body. In the experiments that we have cited, both nitrogen intake and the fuel value of the food per day were lower than in Munk’s experiments, but the utilization of fat and proteid was not sensibly affected. The following tables give the results with ten dogs (including the six dogs already described) for lengths of time ranging from seven to twelve months, the periods indicated being each of ten days’ duration and occurring once each month. In the first table, the utilization of261 fat is shown, the figures given being based on determinations of the amount of fat contained in the excrement. Knowing the amount of fat in the daily food and the amount which passed through the intestine, it is easy to calculate the percentage of fat utilized.
UTILIZATION OF FAT IN PERCENTAGES.
Periods. |
Dogs. |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
1 |
2 |
3 |
4 |
5 |
12 |
13 |
15 |
17 |
20 |
|
1 |
97 |
96 |
93 |
97 |
97 |
96 |
96 |
98 |
98 |
95 |
2 |
96 |
96 |
98 |
98 |
98 |
94 |
95 |
97 |
98 |
95 |
3 |
98 |
97 |
97 |
99 |
96 |
97 |
97 |
98 |
94 |
98 |
4 |
98 |
96 |
97 |
97 |
96 |
94 |
95 |
98 |
97 |
97 |
5 |
96 |
. . |
94 |
98 |
97 |
95 |
95 |
98 |
97 |
96 |
6 |
97 |
98 |
94 |
98 |
97 |
96 |
94 |
97 |
96 |
97 |
7 |
97 |
98 |
98 |
97 |
96 |
93 |
95 |
97 |
98 |
96 |
8 |
. . |
. . |
98 |
96 |
96 |
96 |
93 |
97 |
. . |
. . |
9 |
. . |
. . |
98 |
97 |
98 |
. . |
97 |
98 |
. . |
. . |
10 |
. . |
. . |
98 |
97 |
98 |
. . |
. . |
. . |
. . |
. . |
11 |
. . |
. . |
97 |
92 |
97 |
. . |
. . |
. . |
. . |
. . |
12 |
. . |
. . |
97 |
97 |
. . |
. . |
. . |
. . |
. . |
. . |
It is perfectly plain from these results that there was no falling off in the utilization of fat; the percentage amount digested and absorbed, as in dogs 3 and 4, was just as large at the end of the twelve months’ experiment as at the beginning. Clearly, a so-called low nitrogen intake with dogs does not lead to any loss of power in the utilization of the fat of the food. This being so, it is equally clear that the arguments based on Munk’s results in this direction, and applied to man, are without adequate foundation.
262
UTILIZATION OF NITROGEN IN PERCENTAGES.
| Periods. | Dogs. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
1 |
2 |
3 |
4 |
5 |
12 |
13 |
15 |
17 |
20 |
|
1 |
95 |
91 |
92 |
94 |
91 |
91 |
90 |
93 |
92 |
91 |
2 |
92 |
94 |
94 |
95 |
93 |
90 |
92 |
96 |
92 |
87 |
3 |
91 |
92 |
90 |
91 |
88 |
89 |
86 |
95 |
89 |
91 |
4 |
90 |
85 |
90 |
92 |
91 |
82 |
83 |
91 |
83 |
93 |
5 |
90 |
82 |
88 |
92 |
86 |
85 |
84 |
96 |
91 |
90 |
| 6 | 86 |
87 |
89 |
83 |
86 |
89 |
87 |
94 |
91 |
86 |
7 |
87 |
87 |
90 |
83 |
87 |
83 |
88 |
90 |
93 |
91 |
8 |
. . |
. . |
90 |
83 |
84 |
81 |
89 |
89 |
. . |
. . |
9 |
. . |
. . |
89 |
87 |
92 |
. . |
87 |
89 |
. . |
. . |
10 |
. . |
. . |
93 |
85 |
94 |
. . |
. . |
. . |
. . |
. . |
11 |
. . |
. . |
93 |
81 |
86 |
. . |
. . |
. . |
. . |
. . |
12 |
. . |
. . |
89 |
92 |
. . |
. . |
. . |
. . |
. . |
. . |
The figures in the above table were obtained by determining the amount of nitrogen in the dried excrement from the animals, i. e. the amount that passed through the intestine unchanged;74 and knowing the content of nitrogen in the daily food, the percentage of unabsorbed nitrogen was then easily calculated, after which by simple subtraction the percentage of utilized nitrogen was found. At first glance, it would appear that as the experiments proceeded utilization of nitrogen was less complete. In a sense, this was true, but it was not connected with any impairment of the digestive or absorptive powers of the intestine. It must be remembered that in263 the earlier periods a larger proportion of the ingested nitrogen was in the form of readily digestible meat, but as the latter was reduced in amount larger proportions of vegetable food were introduced in order to maintain the desired fuel value, and consequently the percentage of non-absorbable nitrogen was increased. The well-known difference in the availability of animal and vegetable proteid has already been referred to in other connections; a difference due not so much to any inherent quality in the digestibility of the two forms of proteid as to the presence of cellulose and other material in the vegetable food which retards in some measure the action of the digestive juices. To this cause must be ascribed the slight falling off in the utilization of nitrogen noticeable in most of the experiments. If, however, the figures are compared with those usually obtained on a diet largely vegetable in nature, it will be seen that the utilization of nitrogen by these dogs was in no sense abnormal.
These experiments on the influence of a low proteid diet on dogs, as a type of high proteid consumers, taken in their entirety, afford convincing proof that such animals can live and thrive on amounts of proteid and non-nitrogenous food far below the standards set by Munk and Rosenheim. The deleterious results reported by these investigators were not due to the effects of low proteid or to diminished consumption of non-nitrogenous foods, but are to be ascribed mainly to non-hygienic conditions, or to a lack of care and physiological good sense in the prescription of a narrow dietary not suited to the habits and needs of this class of animals. Further, it is obvious that the more or less broad deductions so frequently drawn from the experiments of Munk and Rosenheim, especially in their application to mankind, are entirely unwarranted and without foundation in fact. Our experiments offer satisfying proof that not only can dogs live on quantities of proteid food per day smaller than these investigators deemed264 necessary, and with a fuel value far below the standard adopted by them; but, in addition, that these animals are quite able on such a diet to gain in body-weight and to lay by nitrogen, thereby indicating that even smaller quantities of food might suffice to meet their true physiological requirements.
The results of these experiments with dogs, which we have recorded in such detail, are in perfect harmony with the conclusions arrived at by our experiments and observations with man, and serve to strengthen the opinion, so many times expressed, that the dietary habits of mankind and the dietary standards based thereon are not always in accord with the true physiological requirements of the body. If these views are correct, and the facts presented seemingly indicate that they are, it is time for enlightened people to give heed to such suggestions, that their lives may be ordered more nearly in accord with the best interests of the body. Physiological economy in nutrition is not a myth, but a reality full of promise for the welfare of the individual and of the community in general. Ignorance on dietary matters should give place to an intelligent comprehension of the body’s needs, and an adequate understanding of how best to meet the legitimate demands of the system for nourishment under given conditions of life. It is said that more than half the earnings of the working people of this country is spent for food. Here, we have suggested another form of economy as worthy of consideration; less important perhaps than that which relates to health and strength, but still calling for thoughtful attention. We cannot afford to be ignorant of these things; we must have definite knowledge of the actual facts, and these can only be obtained by careful research and investigation.
As a prominent writer on nutrition has well said, “The health and strength of all are intimately dependent upon their diet. Yet most people understand very little about what their food contains, how it nourishes them, whether they are economical265 or wasteful in buying and preparing it for use, and whether or not the food they eat is rightly fitted to the demands of their bodies. The result of this ignorance is great waste in the purchase and use of food, loss of money, and injury to health” (Atwater). We all recognize the general force and truth of this statement, but there is a surprising lack of appreciation of the full significance of what is involved thereby. If it is true that the demands of the body for proteid food—which of all foods is the most expensive—are fully met by an amount equal to one-half that ordinarily consumed, and that health and strength are more satisfactorily maintained thereby, it is easy to see how the acquisition of dietary habits leading to consumption of food in harmony with physiological needs will result in a fruitful twofold economy; viz., economy in expenditure, and of still greater moment, economy in the activities of the body by which food and its waste products are cared for.
266
PRACTICAL APPLICATIONS WITH SOME ADDITIONAL DATA
Topics: Proper application of the results of scientific research helpful to mankind. Dietary habits should be brought into conformity with the true needs of the body. The peculiar position of proteid foods emphasized. The evil effects of overeating. What the new dietary standards really involve. The actual amounts of foodstuffs required. Relation of nutritive value to cost of foods. The advantages of simplicity in diet. A sample dietary for a man of 70 kilograms body-weight. A new method of indicating food values. Moderation in the daily dietary leads toward vegetable foods. The experiments of Dr. Neumann. The value of fruits as food. The merits of animal and vegetable proteids considered in relation to the bacterial processes in the intestine. A notable case of simplicity in diet. Intelligent modification of diet to the temporary needs of the body. Diet in summer and winter contrasted. Value of greater protection to the kidneys. Conclusion.
Knowledge has value in proportion to the benefit it confers, directly or indirectly, on the human race. Every new scientific fact or principle brought to light promises help in the understanding of Nature’s laws, and when rightly interpreted and properly applied is sure to aid in the advancement and prosperity of the individual and of the community. Proper methods of living, economical adjustment of the intake to the varying needs of the body, avoidance of excessive waste of foodstuffs and of energy, are all desirable precepts, which rational people presumably are inclined to follow so far as their knowledge and understanding of the subject will permit. Here, as elsewhere, false teaching267 may be exceedingly mischievous and lead to costly errors; while blind reliance upon customs, instinct, and superstitions is hardly in keeping with twentieth-century progress.
Modern scientific methods should give us help in dietetics, as in other branches of hygiene and practical medicine. A few short years ago, diphtheria was a scourge which brought misery to many a home, for there was at hand no adequate means of combating the disease; but scientific research has given us new light, and placed at our command a weapon of inestimable value. Do we hesitate to use it when the occasion arises, because it happens to be out of keeping with old-time customs and traditions? No, we recognize the possibility of help, and as the need is urgent we turn to it quickly, with hope and thankfulness that scientific progress has opened up a pathway of escape from a threatened calamity.
Not many years ago we drank freely of such water as was at hand, without realization of danger from bacteria or disease germs, looking on epidemics of typhoid fever perhaps as a visitation of Divine Providence, in punishment of our many sins and to be borne meekly and with resignation. But all this has changed through the researches of bacteriologists and chemists; scientific facts of the utmost importance have been clearly established; a classification of water-borne diseases has been adopted, and we realize fully that diseases of this order can be kept from our doors by proper precautions applied to our water supply. To-day, epidemics of typhoid fever are traceable solely to the ignorance or carelessness of the individual or of the commonwealth, and the exemption which we of the present generation have from this class of diseases is directly due to the application of precautionary measures based on the information furnished by scientific investigation. It is proper for us to use caution in the acceptance of new ideas, but not that form of caution which refuses change on the ground that what has been is sufficiently good for the present268 and the future. The point of view is ever changing with advance of knowledge, and it is not profitable to exclude opportunities for improvement in personal hygiene and general good health, any more than in other matters that affect the prosperity of the individual or the community.
Dietary habits should be brought into conformity with the true needs of the body. Excessive consumption of proteid food, especially, should be avoided on the ground that it is not only unnecessary and wasteful, but is liable to bring penalties of its own, most undesirable and wholly uncalled for. We may, perhaps, accept these statements at their full value, and yet have a shadow of doubt in our minds as to whether, after all, dietary customs do not harmonize sufficiently at least with true nutritive requirements. All the data that we have presented in the preceding chapters, however, have seemingly given a positive answer to such doubts, and indicate quite clearly that the results of scientific study are opposed to the prevailing dietary standards, especially as regards proteid food. As the celebrated physiologist Bunge has expressed it, “The necessity for a daily consumption of 100 grams of proteid is incomprehensible, so long as we do not know of any function of the body in the performance of which the chemical potential energies of the destroyed proteid are used up.”
Perfectly trustworthy evidence is at hand showing that the needs of the body for potential energy can be fully met, and indeed are more advantageously met, by the non-nitrogenous foods, carbohydrates and fats. The energy of muscle work, as we have seen, comes preferably from the breaking down of non-nitrogenous material, so that there is no special call for proteid in connection with increased muscular activity. In fact, it would appear that the need for proteid food by man is limited to the requirements of growth and development, reinforced by the amount called for in that form of tissue exchange which we have emphasized under the term “endogenous269 proteid metabolism,” or true tissue metabolism. To be sure, there must be a certain reserve of proteid, available in case of emergency, but this is easily established without resorting to excessive feeding.
The peculiar position which proteid foods occupy in man’s dietary naturally make them the central figure, around which the other foods are grouped. No other form of food can take the place of proteid; a certain amount is needed each day to make good the loss of tissue material broken down in endogenous katabolism, and consequently our choice and combination of the varied articles of diet made use of should be regulated by the amount of proteid they contain. But while proteid foods occupy this commanding position, it is not necessary or desirable that they should exceed the other foodstuffs in amount, or indeed approach them in quantity. We must be ever mindful of the fact, so many times expressed, that proteid does not undergo complete oxidation in the body to simple gaseous products like the non-nitrogenous foods, but that there is left behind a residue of non-combustible matter—solid oxidation products—which are not so easily disposed of. In the forceful language of another, “The combustion of proteid within the organism yields a solid ash which must be raked down by the liver and thrown out by the kidneys. Now when this task gets to be over-laborious, the laborers are likely to go on strike. The grate, then, is not properly raked; clinkers form, and slowly the smothered fire glows dull and dies” (Curtis).
Even though no such dire fate overtakes one, the penalties of excessive proteid consumption are found in many ills, for which perhaps the victim seeks in vain a logical explanation; gastro-intestinal disturbance, indigestion, intestinal toxæmia, liver troubles, bilious attacks, gout, rheumatism, to say nothing of many other ailments, some more and some less serious, are associated with the habitual overeating of proteid food. But270 excessive food consumption is by no means confined to the proteid foodstuffs; general overfeeding is a widespread evil, the marks of which are to be detected on all sides, and in no uncertain fashion. One of the most common signs of excessive food consumption is the tendency toward obesity, a condition which is distinctly undesirable and may prove decidedly injurious. Undue accumulation of fat is not only a mechanical obstacle to the proper activity of the body as a whole, but it interferes with the freedom of movement of such muscular organs as the heart and stomach, thereby interposing obstacles to the normal action of these structures. Further, whenever undue fat formation is going on in the body, there is the ever present danger that the lifeless fat may replace the living protoplasm of the tissue cells and so give rise to a condition known as “fatty degeneration.” While a superabundance of fat in the body is a sure telltale of overeating, the absence of obesity is by no means an indication that excess of food is being avoided. There is here, in man as in animal kind, much idiosyncrasy; some persons, especially those endowed with a long and large frame, tend to keep thin even though they eat excessively, while others grow fat much more readily. As a well-known physician has expressed it, “In the one case, the subject burns, instantly and mercilessly, every stick of fuel delivered at his door, whether or not he needs the resulting hot fire roaring within, while the other, miser-like, hoards the rest in vast piles, filling the house from cellar to garret.”
Temperance in diet, like temperance in other matters, leads to good results, and our physiological evidence points out plainly, like a signpost all can read, that there is no demand on the part of the body for such quantities of food as custom and habit call for. Healthfulness and longevity are the prizes awarded for the successful pursuance of a temperate life, modelled in conformity with Nature’s laws. Intemperance, on271 the other hand, in diet as in other matters, is equally liable to be followed by disaster. A physician of many years’ experience, with opportunities for observation among different classes of people, has written, “that overeating tends to shrink the span of life in proportion as it expands the liver is demonstrable both directly and indirectly. Let any actuary of life-insurance be asked his experience with heavy-weight risks, where the waist measures more than the chest, and the long-drawn face of the businessman, at memory of lost dollars, will make answer without need of words. Then let be noted the physique of the blessed ones that attain to green old age, and, in nine cases out of ten, spry old boys—no disparagement, but all honor in the phrase—will be found to be modelled after the type of octogenarian Bryant or nonogenarian Bancroft—the whitefaced, wiry, and spare, as contrasted with the red-faced, the pursy, and the stout. It is true, as has already been mentioned, that in old age much of an adventitious obesity is absorbed and disappears, but the Bryant-Bancroft type is that of a subject who never has been fat at all. And just such is preëminently the type that rides easily past the fourscore mark, reins well in hand, and good for many another lap in the race of life.”75
With these thoughts before us, we may consider briefly just what is involved in these new dietary standards that aim to conform more closely with actual body needs. Referring at first to proteid food, it may be wise to again emphasize the fact that the weight of the body, i. e., the weight of the proteid-containing tissues, as contrasted with excessive fat accumulation, is one of the important factors not to be overlooked when determining the dietary needs of a given individual. As must be perfectly clear, from all that has been said, the man of 170 pounds’ body-weight has more proteid tissue to272 nourish than the man of 130 pounds’ weight, and consequently what will satisfy the requirements of the latter individual will not suffice for the former. We must understand distinctly that no general statement can be made applicable to mankind at large, but due consideration must be given to the size and weight of the individual structure. We have found that the average need for proteid food by adults is fully met by a daily metabolism equal to an exchange of 0.12 gram of nitrogen per kilogram of body-weight. This means a katabolism of three-fourths of a gram of proteid matter daily, per kilogram.
Remembering, however, that the intake of proteid food must be somewhat in excess of the actual proteid katabolism, since not all of the proteid of the food is available, and as this is a variable amount depending upon the proportion of animal and vegetable foods with their different degrees of digestibility and availability, we may place the required intake of proteid at 0.85 gram per kilogram of body-weight, still keeping to maximum figures for safety’s sake. Hence, for a man weighing 70 kilograms or 154 pounds, there would be required daily 59.5 grams—say 60 grams—of proteid food to meet the needs of the body. These are perfectly trustworthy figures, with a reasonable margin of safety, and carrying perfect assurance of being really more than sufficient to meet the true wants of the body; adequate to supply all physiological demands for reserve proteid, and able to cope with the erratic requirements of personal idiosyncrasies. It will be observed that such an intake of proteid food daily is equal to one-half the Voit standard for a man of this weight, while it is still further below the Atwater standard and far below the common practices of the majority of mankind in Europe and America, as indicated by the published dietary studies.
It may not be out of place to state at this point that in the writer’s opinion the use of the terms “standard diet” and “dietary standards,” etc., is objectionable, since such usage273 seems to demand a certain degree of definiteness in the daily diet for which there is no justification. As in the use of the term “normal diet,” there is danger of misinterpretation, and of the assumption that dietary habits should be regulated strictly in accord with certain set principles. This I believe to be altogether wrong; there should be, on the contrary, full latitude for individual freedom, but freedom governed by an intelligence that appreciates the significance of scientific fact and is willing to mould custom and habit into accord with them. What is needed to-day is not so much an acceptance of the view that man requires daily 0.85 gram of proteid per kilogram of body-weight, as a full appreciation of the general principle, which our definite figures have helped to work out, that the requirements of the body for proteid food are far below the customary habits of mankind, and that there is both economy and gain in various directions to be derived by following the general precepts which this view leads to. In other words, there is no advantage, but, on the contrary, much bother and worriment, in attempting to follow out in practice the details of our more or less exact physiological experiments.
The general teaching which they afford, however, can be adopted and put in practice in our daily lives, without striving to follow too closely the so-called standards which our experiments have led to. Again, the sample dietaries adopted in our experiments have no special virtue, aside from the general principle they teach that simple foods are quite adequate for the nourishment of the body, and that the amount of nitrogen or proteid they contain was sufficient to meet the demands of the particular individuals consuming it. Broadening intelligence on matters of food composition is called for on all sides, and as this is acquired together with due appreciation of the relative nutritive values of proteid, fat, and carbohydrate, there is placed at our command the power of intelligent discrimination, with the ability to apply the principles274 set forth in our own way, in harmony with personal likes and dislikes.
To the majority of us, not very familiar with the percentage composition of ordinary food materials, and unaccustomed to the weighing of food in grams, the figures given from time to time may have failed to convey a very definite impression regarding the actual amounts of the various foods made use of. Further, our ideas concerning the bulk of many of the common articles of food necessary to furnish the 60 grams of proteid required daily by a man of 70 kilograms body-weight may be somewhat hazy. The following table, however, will be of service in this direction:
SIXTY GRAMS OF PROTEID ARE CONTAINED IN
Fuel Value* |
||
| One-half pound fresh lean beef, loin | 308 |
calories |
| Nine hens’ eggs | 720 |
|
| Four-fifths pound sweetbread | 660 |
|
| Three-fourths pound fresh liver | 432 |
|
| Seven-eighths pound lean smoked bacon | 1820 |
|
| Three-fourths pound halibut steak | 423 |
|
| One-half pound salt codfish, boneless | 245 |
|
| Two-and one-fifth pounds oysters, solid | 506 |
|
| One-half pound American pale cheese | 1027 |
|
| Four pounds whole milk (two quarts) | 1300 |
|
| Five-sixths pound uncooked oatmeal | 1550 |
|
| One and one-fourth pounds shredded wheat | 2125 |
|
| One pound uncooked macaroni | 1665 |
|
| One and one-third pounds white wheat bread | 1520 |
|
| One and one-fourth pounds crackers | 2381 |
|
| One and two-thirds pounds flaked rice | 2807 |
|
| Three-fifths pound dried beans | 963 |
|
| One and seven-eighths pounds baked beans | 1125 |
|
| One-half pound dried peas | 827 |
|
| One and eleven-twelfths pounds potato chips | 5128 |
|
| Two-thirds pound almonds | 2020 |
|
| Two-fifths pound pine nuts, pignolias | 1138 275 |
|
| One and two-fifths pounds peanuts | 3584 |
|
| Ten pounds bananas, edible portion | 4600 |
|
| Ten pounds grapes | 4500 |
|
| Eleven pounds lettuce | 990 |
|
| Fifteen pounds prunes | 5550 |
|
| Thirty-three pounds apples | 9570 |
|
* Fuel value of the quantity needed to furnish the sixty grams of proteid.
The figures in this table are instructive in many ways. First, it is to be noted that the daily proteid requirement of sixty grams can be obtained from one-half pound of lean meat (uncooked), of which the loin steak is a type. Subject to some variations in content of water, an equivalent weight of lean flesh of any variety, lamb, veal, poultry, etc., will furnish approximately the same amount of proteid. With fish, such as halibut steak, and with liver, three-quarters of a pound are required; while with sweetbreads, four-fifths of a pound are needed to furnish the requisite amount of proteid. Of salt codfish, one-half pound will provide the same amount of proteid as an equivalent weight of fresh beef; while with lean smoked bacon the amount rises to seven-eighths of a pound. Among the vegetable products, it is to be observed that dried peas and beans, almonds and pine nuts, are as rich in proteid as the above-mentioned animal foods, essentially the same weights being called for to provide the daily requirement of proteid. The same is true of cheese, the variety designated having such a composition that one-half pound is the equivalent, so far as the content of proteid is concerned, of a like amount of fresh beef. We must not be unmindful of the fact previously mentioned, however, that there are differences in digestibility among these various foodstuffs which tend to lower somewhat the availability of the vegetable products, also of the cheese, thereby necessitating a slight increase in the amount of these foods required to equal the value to the body of lean meat.
Secondly, passing to the other extreme in our list, we find276 indicated types of foods exceedingly poor in proteid, such as the fruits; notably, bananas, grapes, prunes, apples, etc., also lettuce, and in less degree potatoes. These are the kinds of food that may legitimately attract by their palatability, but do not add materially to our intake of proteid even when consumed in relatively large amounts. Thirdly, we see clearly indicated a radical difference between the animal foods and those of vegetable origin, in that with the former the fuel value of the quantity necessary to furnish the sixty grams of proteid is very small, as compared with a like amount of the average vegetable product. One-half pound of lean meat, for example, with its 60 grams of proteid, has a fuel value of only 308 calories, while two-thirds of a pound of almonds has a fuel value of 2020 calories, and one-half pound of dried peas 827 calories. Naturally, this is mainly a question of the proportion of fat or oil present. With fat meat, as in bacon, the calorific value rises in proportion to increase in the amount of fat, the proteid decreasing in greater or less measure.
The main point to be emphasized in this connection, however, is that a high proteid animal food, like lean meat, eggs, fish, etc., obviously cannot alone serve as an advantageous food for man. We see at once the philosophy of a mixed diet. Let us assume that our average man of 70 kilograms body-weight needs daily 2800 calories. On this assumption, if he were to depend entirely upon lean beef for his sustenance, he would require daily four and a half pounds of such meat, which amount would furnish nine times the quantity of proteid needed by his system. The same would be more or less true of other kindred animal products. On the other hand, certain vegetable foods on our list, such as flaked rice, crackers, and shredded wheat, contain proteid, with carbohydrate and fat, in such proportion that the energy requirement would be met essentially by the same quantity as277 served to furnish the necessary proteid. Passing to the other extreme among the vegetable products, as in potatoes and bananas, for example, we find fuel value predominating largely over proteid content. The ideal diet, however, is found in a judicious admixture of foodstuffs of both animal and vegetable origin. Wheat bread, reinforced by a little butter or fat bacon to add to its calorific value, shredded wheat with rich cream, crackers with cheese, bread and milk, eggs with bacon, meat with potatoes, etc.; the common, every-day household admixtures, provide combinations which can easily be made to accord with true physiological requirements. The same may be equally true of the more complicated dishes evolved by the high art of modern cookery.
Lastly, our table throws light upon certain questions of household economy. The cost of foods is regulated mainly not by the value of the nutrients contained therein, but by other factors of quite a different nature. Relationship between supply and demand naturally counts here as in other directions, but our demand is liable to be based not upon food values, but rather upon delicacy of flavor, palatability, and other kindred fancies, some real and some imaginary. Ordinary crackers can be purchased for ten cents a pound, but if we desire a little stronger flavor of salt and a special box to hold them, we pay eighteen cents a pound. Rolled very thin and thus made more delicate, they cost twenty-five cents, while sold under a special name and perhaps tied with a blue ribbon they cost thirty-five cents a pound. Their nutritive value per pound is the same in all cases, but we pay something for the increased labor of preparation and a good deal for the added attractiveness to eye and palate. We pay twenty-two cents a pound for round steak, thirty-two cents for loin steak, and seventy-five cents a pound for sweetbreads, the high price of the latter being regulated by the relative scarcity of the article and not by its food value. As278 our table indicates, the real value of sweetbread as a source of proteid is only a little more than half that of lean beef. Its fuel value, however, is somewhat more than that of beef, but a little fat added to the latter will more than compensate and at a trifling cost. When we can afford it, we pay the increased price for sweetbreads simply because their delicacy and flavor are attractive to us. We should not do it under the mistaken idea that we are indulging in a highly nutritive article of food, for as a matter of fact it is not only less nutritive than a corresponding weight of lean beef, but in addition it possesses certain qualities, in its high purin-content, that are a menace to good health if indulged in too freely.
Where expense must be carefully guarded, or where the condition of the family purse is such that conflicting demands must be intelligently considered in order to insure wise expenditure and the greatest permanent good of the many, it is well to remember that price is no guarantee whatever of real nutritive value. Two quarts of milk will furnish half the daily fuel requirement of our average man and the entire proteid requirement, while its cost is only sixteen cents. Reinforced by a pound loaf of wheat bread, the energy requirement for the day would be fully met, with surplus nitrogen to store up for future needs, and at an additional cost of only ten cents. A mixture in this proportion, however, would not be strictly physiological, since it is wasteful of proteid, but it may serve to illustrate the point. A better illustration is found in an admixture, quite adequate to supply the daily needs of our average man, both for proteid and energy, composed of one-quarter of a pound of lean beef, two-thirds of a pound of bread, and half a pound of butter, and at a total cost not to exceed thirty cents. The contrast of such prices with what is so commonly paid for table delicacies is somewhat striking; it could be made still more so by drawing upon many common vegetable foods, rich alike in proteid and279 in fuel value, the cost of which is even less than the simple food mixtures just referred to. It is not necessary, however, to enlarge upon this question; it is sufficient to merely emphasize the fact that the exaggerated demand of our present generation for dietetic luxuries is leading us far away from the proverbially simple life of our forefathers, and without adding in any way to the effectiveness of the daily diet. On the contrary, it is in part responsible for the high proteid consumption of the present day, with its attendant evils, and involves a large and unnecessary expenditure without adequate return. The wants of the body for food are far more advantageously met by a simple dietary, moderate in amount and at an expense comparatively slight.
A recent writer,76 in the “British Medical Journal,” a practitioner of medicine in the Highlands of Scotland, has said that these are “facts of common experience in the Highlands of Scotland, and probably among the peasantry of other countries also, where the old beliefs and customs have not too readily given way to the luxuries of civilization. Oatmeal in one form or another is a daily ingredient in the diet of a Highland peasant. The potato also is a staple food, and is consumed in large quantities with salt herring or other fish, and perhaps in some cases salt mutton or pork. Milk and eggs are used by most. The growing consumption of tea, however, and the increasing relish for sweets, candy, pastry, and biscuits, threaten to destroy the old way of living. A typical day’s diet for a crofter or fisherman who still believes in the traditional diet would be somewhat like this:
Breakfast.—Oatmeal porridge or brose with milk; bread, butter, and tea.
Dinner.—Potatoes galore and herrings, or other salt fish.
Supper.—Porridge and milk, or oat bread and cheese, and tea.
“I have often been assured by shepherds that they could work all day ‘on the hill’ after a breakfast of oatmeal brose280 and milk, without fatigue and without feeling hungry, returning in the evening to partake of a dish of broth, potatoes, and salt mutton. In these diets, proteid forms a very small proportion, and yet a hardier race than these shepherds and fishermen cannot be found.” It should be added that “brose” consists of a few handfuls of oatmeal, to which is added boiling water, the mixture being stirred vigorously and placed for a few minutes near the fire. It is then eaten with milk, or better, with cream. In the absence of positive data, it can only be asserted that the above dietary stands for simplicity and frugality. Its proteid-content may be low, but the amount of proteid taken per day by these Highlanders will obviously depend upon the quantity of food consumed. Oatmeal is fairly rich in proteid, and it is quite conceivable that the amount eaten daily may be such as to result in a high proteid exchange.
It will be profitable for us to gain, if possible, a fairly clear idea of the quantities of food requisite for our average man of 70 kilograms body-weight; i. e., the amounts necessary to provide 60 grams of proteid and 2800 calories. With this end in view, we may outline a simple dietary, expressed in terms that will convey a clear impression, showing what may be eaten without overstepping the required limits of proteid or total calories:
BREAKFAST |
|||
Proteid |
Calories |
||
| One shredded wheat biscuit 30 grams |
3.15 |
grams | 106 |
| One teacup of cream 120 grams |
3.12 |
206 |
|
| One German water roll 57 grams |
5.07 |
165 |
|
| Two one-inch cubes of butter 38 grams |
0.38 |
284 |
|
| Three-fourths cup of coffee 100 grams |
0.26 |
. . . 281 |
|
| One-fourth teacup of cream 30 grams |
0.78 |
51 |
|
| One lump of sugar 10 grams |
. . . |
88 |
|
12.76 |
850 |
||
LUNCH |
|||
Proteid |
Calories |
||
| One teacup homemade chicken soup 144 grams |
5.25 |
grams | 60 |
| One Parker-house roll 38 grams |
3.38 |
110 |
|
| Two one-inch cubes of butter 38 grams |
0.38 |
284 |
|
| One slice lean bacon 10 grams |
2.14 |
65 |
|
| One small baked potato 2 ounces, 60 grams |
1.53 |
55 |
|
| One rice croquette 90 grams |
3.42 |
150 |
|
| Two ounces maple syrup 60 grams |
. . . |
166 |
|
| One cup of tea with one slice lemon | . . . |
. . |
|
| One lump of sugar 10 grams |
. . . |
38 |
|
16.10 |
928 |
||
DINNER |
|||
Proteid |
Calories |
||
| One teacup cream of corn soup 130 grams |
3.25 |
grams | 72 |
| One Parker-house roll 38 grams |
3.38 |
110 |
|
| One-inch cube of butter 19 grams |
0.19 |
142 |
|
| One small lamb chop, broiled lean meat, 30 grams |
8.51 |
92 |
|
| One teacup of mashed potato 167 grams |
3.34 |
175 |
|
| Apple-celery lettuce salad with mayonnaise dressing 50 grams |
0.62 |
75 282 |
|
| One Boston cracker, split 2 inches diameter, 12 grams |
1.32 |
47 |
|
| One-half inch cube American cheese 12 grams |
3.35 |
50 |
|
| One-half teacup of bread pudding 85 grams |
5.25 |
150 |
|
| One demi-tasse coffee | . . . |
. . |
|
| One lump of sugar 10 grams |
. . . |
38 |
|
29.21 |
951 |
BREAKFAST |
|||
Proteid |
Calories |
||
| One shredded wheat biscuit 30 grams |
3.15 |
grams | 106 |
| One teacup of cream 120 grams |
3.12 |
206 |
|
| One German water roll 57 grams |
5.07 |
165 |
|
| Two one-inch cubes of butter 38 grams |
0.38 |
284 |
|
| Three-fourths cup of coffee 100 grams |
0.26 |
. . . |
|
| One-fourth teacup of cream 30 grams |
0.78 |
51 |
|
| One lump of sugar 10 grams |
. . . |
88 |
|
12.76 |
850 |
||
LUNCH |
|||
Proteid |
Calories |
||
| One teacup homemade chicken soup 144 grams |
5.25 |
grams | 60 |
| One Parker-house roll 38 grams |
3.38 |
110 |
|
| Two one-inch cubes of butter 38 grams |
0.38 |
284 |
|
| One slice lean bacon 10 grams |
2.14 |
65 |
|
| One small baked potato 2 ounces, 60 grams |
1.53 |
55 |
|
| One rice croquette 90 grams |
3.42 |
150 |
|
| Two ounces maple syrup 60 grams |
. . . |
166 |
|
| One cup of tea with one slice lemon | . . . |
. . |
|
| One lump of sugar 10 grams |
. . . |
38 |
|
16.10 |
928 |
DINNER |
|||
Proteid |
Calories |
||
| One teacup cream of corn soup 130 grams |
3.25 |
grams | 72 |
| One Parker-house roll 38 grams |
3.38 |
110 |
|
| One-inch cube of butter 19 grams |
0.19 |
142 |
|
| One small lamb chop, broiled lean meat, 30 grams |
8.51 |
92 |
|
| One teacup of mashed potato 167 grams |
3.34 |
175 |
|
| Apple-celery lettuce salad with mayonnaise dressing 50 grams |
0.62 |
75 |
|
| One Boston cracker, split 2 inches diameter, 12 grams |
1.32 |
47 |
|
| One-half inch cube American cheese 12 grams |
3.35 |
50 |
|
| One-half teacup of bread pudding 85 grams |
5.25 |
150 |
|
| One demi-tasse coffee | . . . |
. . |
|
| One lump of sugar 10 grams |
. . . |
38 |
|
29.21 |
951 |
The grand totals for the day, with this dietary, amount to 58.07 grams of proteid and 2729 calories. It is of course understood that these figures are to be considered as only approximately correct, but the illustration will suffice, perhaps, to give a clearer understanding of the actual quantities of food involved in a daily ration approaching the requirements for a man of 70 kilograms body-weight. Further, there may be suggested by the figures given for proteid and fuel value of the different quantities of foods, a clearer conception of how much given dietary articles count for in swelling the total values of a day’s intake. Moreover, it is easy to see how the diet can be added to or modified in a given direction. If a little more proteid is desired without changing materially the fuel value of the food a boiled egg can be added to the breakfast, for example. An average-sized egg (of 53 grams) contains 6.9 grams of proteid, while it will increase the fuel value of the food by only 80 calories. Or, if more vegetable proteid is wished for, a soup of split-peas can be introduced, without changing in any degree the calorific value of the diet. Thus, one teacup of split-pea soup (144 grams) contains 8.64 grams of proteid, while the fuel value of this quantity may be only 94 calories. The addition of one banana (160 grams) will increase fuel value 153 calories, but will add only 2.28 grams283 of proteid. If it is desired to increase fuel value without change in the proteid-content of the food, recourse can always be had to butter, fat of meat, additional oil in salads, or to syrup and sugar.
Such a menu as is roughly outlined, however, has perhaps special value in emphasizing how largely the proteid intake is increased by foods other than meats, and which are not conspicuously rich in proteid matter. All wheat products, for example, while abounding in starch, still show a large proportion of proteid. Thus, shredded wheat biscuit (1 ounce), which is a type of many kindred wheat preparations, from bread and biscuit to the various so-called breakfast foods, yields about 3 grams of proteid per ounce and approximately 100 calories. Even potato, which is conspicuously a carbohydrate food owing to its large content of starch, yields of nitrogen the equivalent of at least three-fourths of a gram of proteid per ounce. If larger volume is desired without much increase in real food value, there are always available green foods, such as lettuce, celery, greens of various sorts, fruits, such as apples, grapes, oranges, etc. Too great reliance on meats as a type of concentrated food, on the other hand, augments largely the intake of proteid, and adds a relatively small amount to the fuel value of the day’s ration.
An ingenious method of indicating food values, which promises to be of service in sanatoria and under other conditions where it is desirable to record or correct the diet of a large number of persons, has been devised recently by Professor Fisher.77 The method aims to save labor, and is likewise designed to visualize the magnitude and proportions of the diet. The food is measured by calories instead of by weight, a “standard portion” of 100 large calories being the unit made use of. In carrying out the method, foods are284 served at table in “standard portions,” or multiples thereof. In the words of Fisher, the amount of milk served, for example, “instead of being a whole number of ounces, should be 4.9 ounces—the amount that contains 100 calories. This ‘standard portion’ constitutes about two-thirds of an ordinary glass of milk. Of the 100 calories which it contains 19 will be in the form of proteid, 52 in fat, and 29 in carbohydrate.” In the carrying out of this plan, it is evident that the weight of any food yielding 100 calories becomes a measure of the degree of concentration. From the standpoint of fuel value, olive oil is probably one of the most concentrated of foods, approximately one-third of an ounce containing 100 calories. The following table, taken from Fisher’s description of his method, will serve to show the amounts of several foods constituting a “standard portion,” and also the number of calories in the form of proteid, fat, and carbohydrate:
| Name of Food and “Portion” roughly estimated. |
Weight contain- ing 100 Calories. |
Proteid. | Fat. | Carbo- hydrate. |
Total. | |
|---|---|---|---|---|---|---|
| ounces | grams | calories | calories | calories | calories | |
| Almonds, a dozen | 0.53 |
15 |
13.0 |
77.0 |
10 |
100 |
| Bananas, one large | 3.50 |
98 |
5.0 |
5.0 |
90 |
100 |
| Bread, a large slice | 1.30 |
37 |
13.0 |
6.0 |
81 |
100 |
| Butter, an ordinary pat | 0.44 |
13 |
0.5 |
99.5 |
. . | 100 |
| Eggs, one large | 2.10 |
60 |
32.0 |
68.0 |
. . |
100 |
| Oysters, a dozen | 6.80 |
190 |
49.0 |
22.0 |
29 |
100 |
| Potatoes, one | 3.60 |
100 |
10.0 |
1.0 |
89 |
100 |
| Whole milk, two-thirds glass | 4.90 |
140 |
19.0 |
52.0 |
29 |
100 |
| Beef sirloin, a small piece | 1.40 |
40 |
31.0 |
69.0 |
. . |
100 |
| Sugar, five teaspoons | 0.86 |
24 |
. . . . |
. . . . |
100 |
100 285 |
Obviously, to make use of the “calories per cent” method a table such as the above, covering all common foodstuffs and showing the weight of each food constituting a standard portion, together with the calories of proteid, fat, and carbohydrate in this portion, is necessary. The chief advantage of the method, however, is that it lends itself readily to geometrical representation and affords an easy means of determining the constituents of combinations of different foods by use of a simple mechanism, for a description of which reference must be made to the original paper.
Any attempt to follow a daily routine which accords with the true needs of the body leads necessarily toward foods derived from the plant kingdom, with the adoption of simple dietary habits, and with greater freedom from the exciting influence of the richer animal foods. There is, however, virtue in a simple dietary that appeals and satisfies, and in so doing testifies to the completeness with which it meets the physiological requirements of the body. A physician,78 writing in the “British Medical Journal,” says: “I determined to give the minimum-of-proteid diet a fair trial in my own case. The result was that I was relieved of a life-long tendency to acid dyspepsia and occasional sick headache; my fitness for work, my appetite and relish for food, were increased, without any diminution, but rather a slight increase, in my weight. My practice extends over a wide area of rough mountainous country involving long journeys on cycle, on foot, driving, and in open boats, in fair and foul weather. The muscular exertion and endurance necessary for the work would seem to require a large proportion of proteid and a generous diet altogether, but since I began to experiment I have suffered less than formerly from fatigue, and seem to eat in all a smaller quantity of food. My diet consists of:
286
Breakfast, 8.30 A.M.—Oatmeal cakes, bread and butter, about 1 cubic inch of cheese or bloater paste, marmalade, and one breakfast cup of tea.
Lunch, 1.30 P.M.—Same as breakfast, with occasionally a boiled egg, and sometimes coffee instead of tea.
Dinner, 7 P.M.—Thick soup containing vegetables, with bread, followed by suet pudding or fruit tart; or vegetable stew, containing 2 or 3 ounces of meat, with boiled potatoes, followed by milk pudding and jam, and occasionally a cup of black coffee.”
This statement of personal experience is in close accord with statements that have come to the writer in hundreds of letters during the past two or three years, from persons who have for some reason chosen to follow a more abstemious mode of life. Such testimony has a certain measure of value in that it offers corroborative evidence of the beneficial effects of a moderate diet, more closely in accord with the actual demands of the body for food. It does not, however, carry quite that degree of assurance that scientific evidence, gathered by careful observers and controlled by weights and measures that hold the imagination in check, affords; and so we may turn to a different type of testimony, presented in an elaborate research by Dr. Neumann,79 of the Hygienic Institute at Kiel, an experiment on himself extending through a total of 746 days.
The experiment was divided into three periods. In the first period of ten months the subject, with a body-weight of 66.5 kilograms, consumed daily on an average the amounts of food indicated in the following table. In this same table are also included the daily values, based on the preceding data, for a body-weight of 70 kilograms. Thirdly, the table likewise shows the amounts of utilizable food contained in the foodstuffs actually eaten, on the basis of 70 kilos body-weight.
287
AVERAGE DAILY FOOD FOR TEN MONTHS
Actually consumed by the Subject, 66.5 Kilos |
Calculated for a Body-weight of 70 Kilos |
Utilizable Food for a Body-weight of 70 Kilos |
|
|---|---|---|---|
| Proteid | 66.1 grams |
69.1 grams |
57.3 grams |
| Fat | 83.5 grams |
90.2 grams |
81.2 grams |
| Carbohydrate | 230.0 grams |
242.0 grams |
225.0 grams |
| Alcohol | 43.7 grams |
45.6 grams |
41.0 grams |
| Fuel value | 2309 calories |
2427 calories |
2199 calories |
During this period of ten months, the body-weight of the subject remained practically constant, or indeed showed a slight gain up to 67 kilograms. All the functions of the body, and the general condition of good health, were in no wise impaired; so that in the words of the subject, the amount of food eaten must have been sufficient for the needs of the body. Somewhat striking is the fact that of the 2309 calories in the daily food, more than one-fourth was derived from the beer consumed daily (1200 c.c.). Also noticeable is the relatively small amount of carbohydrate taken daily, only about one-half the quantity designated by Voit as the average requirement of German laborers. Finally, it is to be observed that during this period of ten months, the daily consumption of food as calculated for a man of 70 kilograms body-weight, based on the actual food consumption of the subject with a weight of 66.5 kilos, was not widely different from our own statement of 60 grams of proteid and 2800 calories. The tendency, however, in Dr. Neumann’s experiment was toward lower fuel values and somewhat higher proteid consumption.
In a second period of 50 days, with a slightly larger daily intake, Dr. Neumann observed that his body was laying by nitrogen, i. e., storing up proteid on a daily diet of 76.5 grams288 of proteid and with sufficient fat and carbohydrate to furnish a total fuel value of 2658 calories. In the final period of 8 months, the following data were obtained:
AVERAGE DAILY FOOD FOR EIGHT MONTHS
Actually consumed by the Subject, 71.5 Kilos. |
Calculated for a Body-Weight of 70 Kilos. |
Utilizable Food for a Body-Weight of 70 Kilos. |
|
|---|---|---|---|
| Proteid | 76.2 grams |
74.0 grams |
61.4 grams |
| Fat | 109.0 grams |
106.1 grams |
95.5 grams |
| Carbohydrate | 168.9 grams |
164.2 grams |
152.7 grams |
| Alcohol | 5.5 grams |
5.3 grams |
4.7 grams |
| Fuel value | 2057 calories |
1999 calories |
1766 calories |
During this period, it is to be noted that the fuel value of the day’s food averaged only 2057 calories, which for a body-weight of 70 kilograms would amount to less than 2000 calories. The proteid consumption, however, was larger than we have found to be necessary for a man of the above weight. Still, the facts are in harmony with the general principle that there is no necessity for a daily intake of food such as common usage dictates, there being obviously a wide difference between a minimal daily consumption of 118 grams of proteid and 3000 or more calories, such as is assumed to be needed by a man of 70 kilos, and 74 grams of proteid with 1999 calories. Under the latter conditions, the subject gained a kilogram in weight during the eight months, while the establishment of nitrogen equilibrium testifies to the now generally accepted view that it is quite possible for the body to establish nitrogen equilibrium at different levels, i. e., with different quantities of proteid food and different fuel values.
The diet made use of by Neumann was a mixed one, containing289 a great variety of animal and vegetable foods, but withal simple and moderate in quantity. Calculated per kilogram of body-weight, the average consumption of food material per day during the three periods was as indicated in the following table:
DAILY FOOD CONSUMPTION PER KILOGRAM OF WEIGHT
Proteid. |
Fat. |
Carbohydrate. |
Alcohol. |
Calories |
|
|---|---|---|---|---|---|
grams |
grams |
grams |
grams |
||
| First Period | 0.99 |
1.3 |
34.5 |
0.56 |
34.7 |
| Second Period | 1.10 |
2.3 |
33.4 |
. . |
59.7 |
| Third Period | 1.00 |
1.5 |
23.4 |
0.07 |
28.5 |
The average of daily food consumption for the total of 746 days was as follows: 74.2 grams proteid, 117 grams fat, 213 grams carbohydrate, and 2367 calories. On such a diet, during this long period, equilibrium was satisfactorily maintained, thereby furnishing additional evidence that quantities of food way below the so-called normal amounts are quite adequate to meet the needs of the body. There is no conflict whatever between these results and our own; they both point in the same general direction. Perhaps the one thing that needs to be again emphasized, however, in view of the low fuel values used by Neumann, is that while they proved quite adequate in his case, the demand in this direction is governed largely by the degree of bodily activity. In fact, Neumann’s results with fuel values are in perfect harmony with the data obtained by us with professional men, but the writer is inclined to believe that for the majority of mankind, with the varying degrees of activity and muscular exertion called for, a somewhat larger number of heat units is desirable, and indeed on many occasions demanded.
Still, it is perfectly obvious that custom has greatly exaggerated290 the fuel values required in ordinary muscular work, and such results as are here presented tend to emphasize the true relationship between actual requirements and fuel intake. Further, it must not be overlooked that the rate of proteid katabolism is governed in large measure by the amount of non-nitrogenous food, and consequently a too narrow margin in the consumption of the latter will obviously result in a higher rate of proteid exchange. We are inclined to the belief that a satisfactory degree of bodily efficiency is more liable to be maintained with a somewhat larger consumption of carbohydrate food, combined with a reduction in proteid food to a level nearer our own figures. It will be observed that the average amount of carbohydrate taken daily by Neumann, during the 746 days, was only 213 grams, while the daily consumption of fat averaged 117 grams. These figures are interesting and instructive in many ways, especially as indicating the ease with which the body accommodates itself to a relatively low intake of proteid food, combined with a small proportion of starches and sugars. This relationship between carbohydrate and fat might well occur at times as a natural result of personal taste, but as a general rule it is probably better, from the standpoint of digestibility and general availability, for the daily food to contain a larger proportion of carbohydrate.
Under this head, I would lay special stress upon the value to the body of the natural sugars as well as of starch. We are inclined to deprecate the widespread use of candy, especially among children, and there is no doubt that the too lavish use of sugar in such concentrated form does at times do harm; but when eaten as an integral part of the many available fruits its use cannot be too highly lauded, for both young and old. Oranges, grapes, prunes, dates, plums, and bananas are especially to be commended, and in lesser degree peaches, apricots, pears, apples, figs, strawberries, raspberries, and blueberries.291 In all of these fruits, it is the sugar especially that gives food value to the article, while the mild acids and other extractives, together with the water of the fruit, help in other ways in the maintenance of good health. Where personal taste and inclination are favorably disposed, the first six fruits named can be partaken of freely, and the diet of the young, especially, can be advantageously modified by the liberal use of such articles of food.
Of the other fruits, apples when thoroughly ripe are above reproach if properly masticated, but the raw fruit is somewhat indigestible when swallowed in too large pieces, and may cause trouble to a delicate stomach. A baked apple, on the other hand, is both savory and wholesome, and if served with sugar and cream, for example, constitutes a most healthful and satisfying article of food. Peaches, apricots, and strawberries as ripe fruits are likewise exceedingly valuable, but here personal idiosyncrasy frequently comes to the fore, especially with strawberries, and prohibits their free use. The peculiar acidity of these latter fruits is occasionally a source of trouble, which leads to their avoidance; but this is far less liable to happen with people living on a low proteid diet with its greater freedom from purin derivatives, or uric acid antecedents. Further, there is a tendency on the part of some individuals to suffer from acid fermentation with too liberal use of starches and sugar, but as a rule the advantages of ordinary starchy and natural sugar-containing foods cannot be overestimated. It is certainly wise to give them a conspicuous place in the daily dietary and to encourage their use, especially by children.
As has been stated in several connections, a diet which conforms to the true nutritive requirements of the body must necessarily lead toward vegetable foods. In no other satisfactory way can excess of proteid be avoided, and at the same time the proper calorific value be obtained. This, however,292 does not mean vegetarianism, but simply a greater reliance upon foods from the plant kingdom, with a corresponding diminution in the typical animal foods. This raises the question of the possible relation of diet to the bacterial processes of the intestine, knowing, as we do, that the latter are of primary importance in the causation of certain forms of auto-intoxication, etc. Recent studies have indicated that the bacterial flora of carnivorous animals is quite different from that of herbivorous animals, and this being so, it is easy to see how a predominance of vegetable or animal food may modify the bacterial conditions of the intestinal tract in man. Dr. Herter80 has reported the presence in the intestines of cats, dogs, tigers, lion, and wolf of many spore-holding bacilli, as well as free spores and vegetative forms of anærobic organisms; some of which at least are decidedly pathogenic when injected into the subcutaneous connective tissue, leading to serious and even fatal results within twenty-four hours. With herbivorous animals, on the other hand, such as the buffalo, goat, horse, elephant, etc., the predominating organisms are of a different order from those found in the intestines of the carnivora; proving practically non-pathogenic, or only slightly so, when injected subcutaneously, and less disposed to produce putrefactive changes or other chemical decompositions.
In the words of Dr. Herter, “These differences in the appearance and behavior of the bacteria derived from typical carnivora and herbivora suggest that the habit of living upon a diet consisting exclusively of raw meat entails differences in the types of bacteria that characterize the contents of the large intestine. The occurrence of considerable numbers of spore-bearing organisms in the carnivora points to the presence of anærobic putrefactive forms in great numbers.293 The results of subcutaneous inoculations into guinea-pigs bear out this view and indicate that the numbers of organisms capable of producing a hemorrhagic œdema with tissue necrosis, with or without gas-production, are very considerable. . . . The observations recorded are of much interest in relation to the bacterial processes and nutrition of herbivorous as distinguished from carnivorous animals, and are significant furthermore for the interpretation of bacterial conditions found in man. The question arises whether the abundant use of meat over a long period of time may not favor the development of much larger numbers of spore-bearing putrefactive anærobes in the intestinal tract than would be the case were a different type of proteid substituted for meat.” While it may be said truly that observations of this character are as yet not sufficiently numerous or conclusive to warrant positive or sweeping statements, yet there is a suggestion here well worthy of thoughtful consideration in its general bearing on the nutrition of mankind.
Simplicity in diet, with or without complete abstinence from meat, is often resorted to as a means of relief from bodily ailments, and such cases sometimes afford striking illustrations of the adequacy and benefits of a relatively low intake of food. Cases of this sort, perhaps, are more frequently observed among elderly people, where the daily requirements are not so great as with younger and more active persons, but they offer evidence in support of our main thesis that dietary habits are no guarantee of bodily requirements. I have in mind the details of an exceedingly interesting case reported with much care by Dr. Fenger;81 the case of a man who at 61 years of age, after a long period of poor health, brought himself quickly into a condition of sound health by a daily diet characterized by extreme simplicity and with an exceedingly low294 fuel value. The daily diet made use of during the fifteen years the subject was under examination consisted of the following articles:
1889–1892: 1 egg, 1 quart of oatmeal soup, 2 quarts of skim milk, 1 12 ounces of red wine, 14 ounce of sugar.
1892–1894: 2 eggs, 1 quart of oatmeal soup, 2 quarts of skim milk, 1 12 ounces of red wine, 14 ounce of sugar.
1894–1900: 3 eggs, 1 pint of oatmeal soup, 2 quarts of skim milk, 1 12 ounces of red wine, 14 ounce of sugar, 2 ounces of plum and raspberry juice.
1900–1903: 3 eggs, 1 pint of barley soup, 3 pints of sweet milk, 1 pint of buttermilk, 1 12 ounces of red wine, 14 ounce of sugar, 2 ounces of plum and raspberry juice.
It will be observed that during these fifteen years the subject partook of no meat whatever, and further, that the diet was wholly in fluid form. At the close of this long period, the subject, being then 75 years of age, was reported as well and in good health, with satisfactory physical condition for a person of his years. He was a man of small body-weight, only 42 kilograms, but during this period of voluntary restriction in diet, he suffered no loss. It is perhaps worthy of comment also that all through this lengthy period no salt was taken other than what was naturally present in the simple foods made use of. The point to attract our attention especially, however, is that for fifteen years, during which the quality and quantity of this man’s food was carefully observed, body-weight, general good health, and physical vigor were all maintained, together with freedom from the ills of previous years and with a daily diet characterized by extreme simplicity. The chemical composition of the diet was likewise peculiar, particularly in its exceedingly low fuel value. The following table shows the amounts of proteid, fat, and carbohydrate consumed daily during the four periods designated:
295
Period. |
Proteid. |
Fat. |
Carbo- hydrate. |
Calories. |
Calories per Kilo- gram. |
Proteid per Kilo- gram. |
|---|---|---|---|---|---|---|
grams |
grams |
grams |
grams |
|||
1889–1892 |
79.8 |
21.7 |
152.0 |
1125 |
26 |
1.90 |
1892–1894 |
85.2 |
27.0 |
152.0 |
1200 |
28 |
2.03 |
1894–1900 |
87.0 |
30.1 |
150.1 |
1230 |
29 |
2.07 |
1900–1903 |
84.4 |
73.7 |
148.3 |
1600 |
38 |
2.00 |
Especially noticeable here is the low intake of fat and carbohydrate, with the corresponding low fuel value, and also the relatively high consumption of proteid, averaging 2.0 grams daily per kilogram of body-weight. Dr. Fenger concludes that for a man of this age and weight, with the relative inactivity characteristic of old age, a heat value in the intake of 30 calories per kilogram of body-weight is quite sufficient for the needs of the body. This may be quite true, but to maintain nitrogen equilibrium under such conditions requires a larger intake of proteid food than is desirable. It will be observed that in the last period of four years a very decided change in the diet was instituted; proteid was diminished somewhat, but the noticeable change was the decided increase in fat, produced in large measure by the substitution of whole milk, with its contained cream, for skim milk. In the words of Dr. Fenger, this change was necessitated by the appearance of gout in the subject. From superficial examination of the dietary of the preceding eleven years there would seem no occasion for criticising the subject for high living, and yet I believe we are quite within the limits of reason in saying that the proteid exchange for a subject of this body-weight was altogether too high. The heat requirements of the body were being met in an unnecessarily large degree from the breaking down of proteid material, with consequent formation of excessive296 nitrogenous waste, among which uric acid was plainly conspicuous.
One comment to be made here is that meat and other rich purin-containing foodstuffs are not the only source of gout and uric acid. Excessive proteid katabolism, both exogenous and endogenous, is a possible source of danger in this respect, and the above subject, though living on an exceptionally simple diet, was consuming far more proteid per kilogram of body-weight than was necessary or desirable. Increase of fatty food naturally served to diminish the rate of proteid katabolism, and this could have been advantageously accompanied by a still greater reduction in the amount of proteid ingested, and a larger addition of non-nitrogenous foodstuffs. In old age, there is naturally a slowing down of the metabolic processes, and both nitrogen equilibrium and body equilibrium can be satisfactorily maintained by a relatively small intake of food and with gain to the body; but there is every reason to believe that economy in proteid food can be more advantageously adopted than economy in non-nitrogenous foodstuffs.
Finally, we may call attention to the many possibilities of an intelligent modification of the daily diet to the temporary needs of the individual. The season of the year, summer and winter, the climate, the degree of activity of the body, the state of health, temporary ailments, etc., all present special conditions which admit of particular dietetic treatment. In hot summer weather, for example, there is plainly less need for food than in the cold winter season, especially for fat with its high calorific value. During the cold part of the year, the lower temperature of the surrounding air, with the tendency toward greater muscular activity, calls for more extensive chemical decomposition in order to meet the demand for heat, and the energy of muscular contraction. There is perhaps no special reason for any material change in the amount297 of proteid food consumed in the two seasons, except in so far as it may seem desirable at times to take advantage of the well-known stimulating properties of proteid to whip up the general metabolism of the body, in harmony with the principle that all metabolic processes may need spurring to meet the demands of a greatly lowered temperature in the surrounding air.
Fuel value, however, should be increased somewhat during the winter months in our climate. Fat promises the largest amount of energy, but there is more of a tendency to store up excess of fat than of carbohydrate, hence the latter foods have certain advantages as a source of the additional energy needed during cold weather. In warm weather, it should be our aim to diminish unnecessary heat production as much as possible, though it must be remembered that the body is to be maintained approximately at least in equilibrium, and this calls for an adequate amount of food. Lighter foods, however, may be advantageously employed, such as fruits, vegetables, fresh fish, etc. Fats and fat meats especially are to be avoided, not only because there is no specific need for them, but particularly on account of a greater sensitiveness of the gastro-intestinal tract during the hot seasons of the year, that is liable to result in a disturbance whenever undue quantity of rich or heavy food is taken. Further, in hot summer weather we may advantageously live more largely on foods served cold, and thereby avoid the heat ordinarily introduced into the body by hot fluids and solids. These, however, are all obvious physiological truths, constituting a form of physiological good sense the application of which calls for no special expert knowledge.
Less obvious, though no less important, is the partial protection that can be afforded to weakened or disabled kidneys by judgment and discrimination in the matter of diet. In acute or chronic nephritis, forms of so-called Bright’s disease,298 is there not danger of overtaxing organs already weakened by placing upon them the daily duty of excreting large amounts of solid nitrogenous waste, as well as of the various inorganic salts which are so intimately associated with many of the organic foodstuffs? The consumption of excessive and unnecessary amounts of proteid food simply means the ultimate formation of just so much more urea, uric acid, etc., which must be passed out through the kidneys. In the words of Bunge, “There is no organ in our body so mercilessly ill treated as the kidneys. The stomach reacts against overloading. The kidneys are obliged to let everything pass through them, and the harm done to them is not felt till it is too late to avoid the evil consequences.” It would seem the part of wisdom, therefore, to adjust the daily intake of proteid food to as low a level as is consistent with the true needs of the body, in those cases where the kidneys are at all enfeebled, or where it seems desirable to exercise due precaution as a possible means of prevention.
Equal care is frequently called for in connection with the mineral matters which enter so largely into many natural foodstuffs, or which are introduced as condiments. As an illustration, we may note one or two peculiarities in the distribution of sodium and potassium salts in the tissues of the body. Potassium is an indispensable constituent of every living cell, and the latter has the power of absorbing and holding on to such amounts of this particular element as may be necessary for the functional activity of the tissue of which it is a part. Sodium, on the other hand, stands in a different relationship to living structures. It is widely distributed, but in the higher animals, as in man, sodium salts are most abundant in the fluids of the body, notably in the plasma of the blood. Herbivorous animals have a strong liking for sodium chloride or common salt, but this is not true of carnivorous animals; indeed, the latter animals have a great299 dislike for salty articles of food. Vegetable products are all rich in potassium salts, whereas ordinary animal foods, such as meat, eggs, milk, and blood, are relatively poor in this element.
It is claimed that the abundance of potassium salts in vegetable foods is the cause of the apparent need for sodium chloride by herbivorous animals, and in lesser degree by man. This is explained by supposing that when the salts of potassium reach the blood by absorption of the vegetable foods, an interchange takes place with the sodium chloride of the blood plasma. “Chloride of potassium and the sodium salt of the acid which was combined with the potassium are formed. Instead of the chloride of sodium, therefore, the blood now contains another sodium salt, which did not form part of the normal composition of the blood, or at any rate not in so large a proportion. A foreign constituent or an excess of a normal constituent, i. e., sodium carbonate, has arisen in the blood. But the kidneys possess the function of maintaining the same composition of the blood, and of thus eliminating every abnormal constituent and any excess of a normal constituent. The sodium salt formed is therefore ejected by the kidneys, together with the chloride of potassium, and the blood becomes poorer in chlorine and sodium. Common salt is therefore withdrawn from the organism by the ingestion of potassium salts. This loss can only be made up from without, and this explains the fact that animals which live on a diet rich in potassium, have a longing for salt” (Bunge). It is certainly a fact worthy of note that man takes only one salt as such in addition to those that are naturally present in his food, and it is equally significant that sodium chloride is by no means lacking in ordinary foodstuffs. If the individual lives entirely on animal foods, he has no desire for salt, but as soon as he adopts a vegetable diet the craving for salt shows itself. Vegetable foods, however, are not all300 alike in their content of potassium salts; some, like rice, contain relatively little, while others, like potatoes, peas, and beans, are comparatively rich in this element.
We may recognize in these statements a physiological demand for a certain amount of salt, especially when vegetable foods enter into the daily dietary, but there is no justification for the employment of such quantities as are generally made use of. Where the vegetable food is largely rice, a small fraction of a gram of salt is really sufficient for all physiological purposes; and in those cases where ordinary cereals, legumes, potatoes, etc., constitute the chief part of the dietary, a few grams of salt, at the most, will suffice to meet the daily needs. Common usage, however, frequently raises the amount consumed to 25 grams or more per day, the bulk of which is at once eliminated through the kidneys; thereby entailing a certain amount of renal activity, which must, it would seem, constitute something of a strain upon organs ordinarily hard worked at the best. “Do we not impose too great a task upon them, and may it not be fraught with serious consequences? When on a diet of meat and bread, without salt, we excrete not more than from 6 to 8 grams of alkaline salts in twenty-four hours. With a diet of potatoes, and a corresponding addition of salt, over 100 grams of alkaline salts pass through the kidneys in the day. May not there be danger in this? The habit of drinking spirituous liquors, which moreover is reckoned one of the causes of chronic nephritis, also brings about the immoderate use of salt, and thus one sin against nature leads to another” (Bunge).
The moral we would draw (from these observations) is that in weakened conditions of the kidneys there is reason in reducing the rate of proteid exchange to the lowest level consistent with the maintenance of equilibrium and the preservation of strength and vigor, thereby diminishing the amount of nitrogenous waste to be eliminated and the consequent strain301 upon these organs. Further, there is suggested moderation in the amount of salt to be used daily, and some circumspection in the amount and quality of vegetable foods consumed in order to regulate more effectually the quantity of saline waste to be handled by the kidneys. These conclusions are just as worthy of consideration as the more obvious rule that in diabetes or glycosuria proper precaution must be observed in the eating of carbohydrate foods. In gout and rheumatism, accumulated physiological knowledge teaches plainly the necessity of avoiding those foods that are rich in purin-containing compounds. Uric acid owes its origin in part at least to substances of this class; and as an ounce of prevention is worth more than a pound of cure, we may by proper moderation in the use of such foods save ourselves from the disagreeable effects of accumulated uric acid deposits.
In conclusion, the nutrition of man, if it is to be carried out by the individual in a manner adapted to obtaining the best results, involves an intelligent appreciation of the needs of the body under different conditions of life, and a willingness to accept and put in practice the principles that scientific research has brought to light, even though such principles stand opposed to old-time traditions and customs. The master words which promise help in the carrying out of an intelligent plan of living are moderation and simplicity; moderation in the amount of food consumed daily, simplicity in the character of the dietary, in harmony with the old saying that man eats to live and not lives to eat. In so doing there is promise of health, strength, and longevity, with increased efficiency, as the reward of obedience to Nature’s laws.
303
Abderhalden, Emil, 35
Absorption, a physiological process, 41
diffusion as a factor in, 41
from the stomach, 31
in intestine, 37
of fats, in dogs on low proteid diet, 233, 261
of food products, by blood, 44
of peptones, 41
of proteid in dogs on low proteid diet, 233, 262
of proteid products, 47
of proteoses, 41
osmosis, as factor in, 41
paths of, 44
reconstruction of proteid during, 42
selective action, of sugars, 47
uric, 73
uric, excretion of, as influenced by diet, 144
Acids, amino, 34
diamino, 34
Adenase, 71
Adenin, 72
Aldehydase, 64
Amylopsin, 32
Anabolism, 50
Animals, influence of low proteid diet on high proteid, 231, 233, 243
Animal starch, see Glycogen
Appetite, in relation to food requirements, 162
Argutinsky, views on muscle work, 123
Assimilation limits of sugars, 47
Athlete, photograph of, 190
Athletes, fuel value of food of, on low proteid diet, 198
strength tests of, on low proteid diet, 206304
true proteid requirement of, 186
Atwater and Benedict, 109, 111
Autodigestion (see Autolysis), 63
Autolysis, 12
Availability, of foods, 12
of carbohydrates, as source of energy, 45
Bacterial flora in intestine, of carnivora, 292
of herbivora, 292
Bacterial processes in intestine, in relation to food, 292
Balance, nutritive, as affected by various factors, 117, 118
Basal energy exchange, 104
Beaumont, William, on movements of stomach, 27
Benedict, F. G., see Atwater and Benedict
Bergell and Lewin, 36
Beriberi, and diet, 224
Blood, absorption of food products by, 44
behavior of disaccharides when introduced into, 39
effects of injection of proteoses and peptones into, 41
relation of sugar in, to glycogen, 46
sugar in, 45
Body, amounts of food required to furnish proteid needs of, 274
efficiency of, as a machine, 111
equilibrium, 78
nature of oxidation in the, 60
needs of nitrogen by, 4
needs for food by, 169
needs and dietary habits, 268
relation of oxygen to decompositions in, 61
resistance, see Resistance
sample dietary supplying needs of, 280
site of oxidation in, 62
surface, relation to energy exchange, 104, 105
surface, relation to nitrogen requirement in dogs, 248
Body-weight, on low proteid diet, 175, 181, 185, 190, 199, 245–255
relation to proteid requirement, 184, 188, 198, 227
Bright’s disease, see Nephritis
Breisacher, L., on minimum proteid requirement, 172
Bunge, 124
Calorie, 14
Calorimeter, respiration, 102
Cane sugar, assimilation limit of, 47
behavior when introduced into blood, 39
utilization of, 40
Cannon, W. B., on muscular movements of stomach, 28, 29305
Carbon dioxide, output in rest, 111, 112
dioxide, output during work, 111, 123
equilibrium, 84
excretion, during fasting, 84
moiety of proteid, 129
Carnivora, bacterial flora in intestine of, 292
Carbohydrates, as food, 6
as fuel, 6
as heat producers, 58
as proteid sparers, 92
as source of energy, 128
as source of energy in fasting, 81
as source of energy in work, 58
availability of, 13
availability of, as source of energy, 45
composition of, 5
formation from proteid, 129
fuel value of, 15
in foodstuffs, 7
liver as regulator of, 45
respiratory quotient of, 107
Casein, cleavage products of, 70
Caspari and Glässner, on minimum proteid requirement in man, 172
Cellulose, in vegetables, influence on digestion, 263
Chemical character of proteid, influence on nutrition, 256
composition of foodstuffs, 7
Circulating proteid, 134
Clapp, S. H. (see Osborne and Clapp, on proteid cleavage products), 258
Cleavage, oxidative, 61
Climbing, oxygen consumption in, 116
Cogan, Thomas, on temperance in food, 166
Cohnheim, Otto, on proteid decomposition, 36
Composition, of proteid, 3
of carbohydrate, 5
of fat, 6
Cornaro, Louis, on temperance in food, 168
Cost of foods in relation to nutritive value, 277
Creatin, 74
Creatinin, 74
excretion, as influenced by diet, 144
Curtis, Edward, Nature and Health, 2, 5, 214
Dapper, Max, 99
Dangers of underfeeding, 214
Degeneration, fatty, 270
Dextrose, 37306
assimilation, limit of, 47
utilization of, 40
Diabetes, phloridzin, 130
Diamino acids, 34
Diet, and beriberi, 224
and renal activity, 297
effects of exclusive proteid, upon rats, 239
effects of intemperance in, 270
effects of rice, on rats, 240
fat absorption in dogs on low proteid, 233, 261
influence of, on creatin in excretion, 144
exclusive proteid, on progeny in rats, 240
on growth in rats, 239
monotony in, 242
on oxygen consumption in man at rest, 126
on oxygen consumption in man at work, 126
on respiratory quotient in man at rest, 126
on respiratory quotient in man at work, 126
rice, on growth in rats, 240
on urea excretion, 144
on uric acid excretion, 144
vegetable, upon dogs, 254, 256
in relation to nephritis, 297
in relation to nitrogen distribution in urine, 144
in relation to seasons of the year, 296
of Highlanders, 279
low proteid, influence on body-weight in dogs, 245, 249, 250, 251, 252, 255
nitrogen excretion during severe work on exclusive proteid, 123, 124
philosophy of a mixed, 92, 276
relation of endurance to low proteid, 210, 212
relation of inorganic salts to, 299, 300
relation of work to, 126
relation of vegetable food to low proteid, 291
sample, of soldiers, 194
sample, in experiments on true proteid requirement in man, 178, 182, 189, 195
simplicity in, advantages of, 279, 293
temperance in, 270
utilization of fat in dogs on low proteid, 261
utilization of nitrogen in dogs on low proteid, 262
Diets, normal, see Standard diets
standard, 155
Dietary habits, in relation to needs of body, 268
of fruitarians, 215
of Japanese, 225
sample, supplying needs of body, 280
standards, use of the term, 272
Dietetic customs of mankind, 154307
Dietetics, habit in, 159
Diffusion, as factor in absorption, 41
Digestibility, see Availability
Digestion, gastric, of proteids, 26
importance of gastric, 30
influence of cellulose in vegetables on, 263
in the stomach, 25
object of gastric, 30
of fat, in intestine, 36
of fat, in stomach, 36
of starch, 21
products of pancreatic, of fats, 36
products of pancreatic, of proteids, 34, 67
products of pancreatic, of starch, 37
products of salivary, 21
salivary, in stomach, 23
Digestive products, reconstruction of proteid from, 42
Disease, relation of excessive proteid consumption to, 269
Dogs, effects of low proteid diet on, 232–236, 245–255
fasting experiments on, 82
fat absorption in, on low proteid diet, 233, 261
fuel value requirement of, 234, 236, 245–255
influence of low proteid diet upon body-weight in, 245–255
influence of vegetable diet on, 254, 256
nitrogen requirement of, 234, 235, 236, 245–255
photographs of, 248
proteid absorption in, on low proteid diet, 233, 262
proteid requirement, experiments by Munk, 232
proteid requirement, experiments by Rosenheim, 234
proteid requirement, experiments by Jägerroos, 236
proteid requirement, experiments by author, 243
utilization of fat in, on low proteid diet, 261
utilization of nitrogen in, on low proteid diet, 262
Disaccharides, utilization of, 40
Edestin, cleavage products of, 70
Efficiency of body, as a machine, 111
Egg albumin, cleavage products of, 70
Endogenous metabolism, 145, 146
Endurance, relation of, to low proteid diet, 210, 212
Energy, availability of carbohydrates, as source of, 45
basal exchange, 104
carbohydrate as source of, 128
carbohydrate as source of, in fasting, 81
conservation of, in man, 103
exchange, effect of muscular work, 109, 110, 113, 115
exchange, factors modifying, 105, 106308
exchange, in relation to work, 119
exchange proportional to body surface, 104, 105
fat as source of, 128
fat as source of, in fasting, 81
foods as source of, 15
metabolism of, in man, 103
of muscle contraction, 121
origin of, in fasting, 81
output, in man, 103
produced by man, 106
proteid as source of, 122, 123, 124, 129
proteid as source of, in fasting, 81
source of, during fasting, in work, 125
Enterokinase, 33
in gastric juice, 25
in pancreatic juice, 32
in saliva, 20
reversible action of, 21
specificity of, 21
Equilibrium, carbon, 84
nitrogenous, 78
of body, 78
Erepsin, 34
Exchange, basal energy, 104
of energy, as affected by work, 109, 110, 113, 115, 119
of energy, factors modifying, 105, 106
of energy, relation to body surface, 104, 105
Exogenous metabolism, 145, 146
Fasting, carbohydrates as source of energy in, 81
excretion of carbon during, 84
excretion of nitrogen during, 80, 82, 84
experiments on dogs, 82
fat as source of energy in, 81
fuel value during, 86
fuel value of fat, metabolized during, 86
metabolism of fat during, 84
nitrogen excretion during, 80, 82, 84
origin of energy in, 81
proteid as source of energy in, 81
proteid metabolism during, 83
relation of nitrogen excretion to work during, 125
source of energy for work during, 125
absorption in dogs on low proteid diet, 233, 261
as food, 6
as fuel, 6
as source of energy, 128
as source of energy during work, 58
as source of energy in fasting, 81
composition of, 6
digestion of, in intestine, 36
digestion of, in stomach, 36
fuel value of, 15
fuel value of, metabolized during fasting, 86
hydrolysis of, 36
influence of feeding, on body fat, 44
in foodstuffs, 7
laying on of, from overfeeding, 98, 99
metabolism during fasting, 84, 86
respiratory quotient of, 107
saponification of, 36
specificity of body, 44
synthesis of, 43
utilization of, in dogs on low proteid diet, 261
Fats, availability of, 13
as heat producers, 58
as proteid sparers, 92
Fatty degeneration, 270
Fatigue, relation to low proteid diet, 208
Fenger, S., 293
Fick and Wislicenus, on source of muscular energy, 121
Fischer, Emil, 21
Fisher, Irving, on endurance and low proteid diet, 210
on method of indicating food values, 283
Folin, Otto, theory of proteid metabolism, 144
Food, absorption and utilization of, in dogs on low proteid diet, 261, 262
amounts, required for proteid needs of body, 274
as fuel, 6
as source of energy, 15
availability of, 12
carbohydrates as, 6
character of, in relation to bacterial processes in intestine, 292
consumption and obesity, 270
consumption, relation to prosperity, 160
fats as, 6
fuel value of, 274
of fruitarians, 217
in experiments on proteid requirement, athletes, 198
in experiments on proteid requirement, professional men, 178, 180, 185
in experiments on proteid requirement, soldiers, 198
fuel value requirement of, in dogs, 234, 236, 245–255310
influence of, on respiratory quotient, 107
needs of body for, 169
of man, 2
real need of body for proteid, 272
relation of appetite to, 162
relation of nutritive value and cost of, 277
requirements, factors modifying, 165
value of fruits as, 290
values of, method of indicating, 283
Foods, respiratory, 58
time, remain in stomach, 29, 30
Foodstuffs, carbohydrate in, 7
composition of, 7
fat in, 7
fuel value of, 7
inorganic salts in, 7
organic, 3
plastic, 58
proteid in, 7
water in, 7
Fritz, photograph of, 199
Fruitarians, dietary of, 215
fuel value of food of, 217
proteid consumption of, 217
Fruits, value of, as food, 290
Fuel, carbohydrate as, 6
fat as, 6
proteid as, 6
Fuel value, in fasting, 86
of carbohydrate, 15
of fat, 15
of fat metabolized during fasting, 86
of food, in experiments on proteid requirement, athletes, 188
of food, in experiments on proteid requirement, professional men, 178, 180, 185
of food, in experiments on proteid requirement, soldiers, 198
of food of fruitarians, 217
of foods, 274
of foodstuffs, 7
of proteid, 15
of proteid metabolized during fasting, 86
requirement in the dog, experiments by Munk, 234
requirement in the dog, experiments by Rosenheim, 236
requirement in the dog, experiments by Jägerroos, 236
requirement in the dog, experiments by author, 245–255311
Gastric digestion, importance of, 30
object of, 30
products of, 26
Gastric juice, action on milk, 26
influence of diet upon flow of, 25
pepsin in, 25
psychical stimulation of, 24
Gastric secretion, 24
Glässner, see Caspari and Glässner
Gliadin, cleavage products of, 70, 259
Glutaminic acid, 34, 67, 70, 259
Glutenin, cleavage products of, 259
Glycerin, 36
Glycocoll, 67
Glycogen, formation from proteid, 130
in liver, 46
relation to sugar of blood, 46
Growth, influence of diet on, in rats, 239
Guanase, 71
Guanin, 72
Habit, in dietetics, 159
Heat, furnished by fats and carbohydrates, 58
production during sleep, 104, 105
production in work, 110
Herbivora, bacterial flora in intestine of, 292
Herter, C. A., on bacterial flora, 292
Hirschfeld, Felix, on minimum proteid requirement, 170
Hofmeister, Franz, on sugar assimilation, 47
Hunt, Reid, on low proteid diet and body resistance, 226
Hunter, Andrew, see Watson and Hunter
Hydrochloric acid, in gastric juice, 25, 26
Hydrolysis, of fats, 36
Hypoxanthin, 72
Indol, 37
Inorganic salts, and renal activity, 298, 300
in foodstuffs, 7
in nutrition, 2
Intemperance in diet, effects of, 270
Intermediary metabolism, see Exogenous metabolism
Intestine, absorption in, 37
chemical changes in, 33
putrefaction in, 37
bacterial flora of, 292
Invertase, 40
Jägerroos, B. H., on proteid requirement in the dog, 236
Japanese Army and Navy, rations of, 224
Japanese, dietary of, 225
fuel value of food of, 219, 221
proteid consumption by, 219, 221
Katabolism, 50
nature of proteid, 75
oxygen in, 62
relation to intracellular enzymes, 75
Klemperer, on proteid requirement, 171
Lactase, 40
Lavoisier, views on oxidation, 56
Leucosin, cleavage products of, 259
Levulose, assimilation limits of, 47
Lewin, see Bergell and Lewin
Liebig, views on oxidation, 57, 120
Lipase, 32
Lipolysis, by pancreatic juice, 36
Liver, function of, as regulator of carbohydrate, 45
glycogen in, 46
synthesis of proteid by, 48
Luxus consumption, of proteid, 59
Lüthje, 101
Lymphatics, absorption of food products by, 44
behavior when introduced into blood, 39
Man, conservation of energy in, 103
energy produced by, 106
experiments on oxygen consumption in, 126
fasting experiments on, 80, 84313
food of, 2
metabolism of energy in, 103
minimum proteid requirement in, 170, 171, 172, 174–208
work experiments on, 110–116
Mastication, importance of, 23
Meat, influence on growth in rats, 239
Metabolic changes as influencing respiratory quotient, 108
Metabolism, 51
and old age, 296
endogenous, 145
exogenous, 145
Folin’s theory of proteid, 144
influence of proteid on, 83
influence of carbohydrates on proteid, 92, 94, 95, 96, 97
influence of fat on proteid, 92, 93, 96, 97
influence of proteid on proteid, 88
of energy in man, 103
oxidation in, 60
of proteid during fasting, 83, 86
Pflüger’s theory of proteid, 138
processes of, 51
significance of exogenous and endogenous proteid, 49
significance of proteid, 131
Voit’s theory of proteid, 134
Methyl glycocoll, see Sarcosin
Methyl guanidin, 74
Milk sugar, assimilation limit of, 47
behavior when introduced into blood, 39
utilization of, 40
Mineral matter, see Inorganic salts
Minimum proteid requirement, 59
Mixed diet, philosophy of a, 92, 276
Monotony of diet, influence of, 242
Morphotic proteid, 134
Munk, Immanuel, on proteid requirement in the dog, 232
Muscular movements of stomach, 27–30
Needs of body for food, 169
Nephritis, in relation to diet, 297
Neumann, R. O., on low proteid diet, 286
Nitrogen, distribution of, in the urine in relation to diet, 144
needs by body, 4
utilization of, in dogs on low proteid diet, 262
Nitrogen excretion, as influenced by proteid, 59, 87, 90
during work in fasting, 125
during excessive work, 114, 127314
during hard work on proteid diet, 123, 124
in experiments on proteid requirement, in dogs, 245, 249, 250, 251, 252, 255
in experiments on true proteid requirement, athletes, 187, 188
in experiments on true proteid requirement, professional men, 176, 177, 181, 185
in experiments on true proteid requirement, soldiers, 199, 200, 201
relation to work, 122, 123, 124
Nitrogen equilibrium, on low proteid diet, 176, 177, 181, 188, 200, 201, 249, 250, 251, 252, 255
Nitrogen requirement, in dogs, 234–236, 245–255
in man, 180, 184, 185, 187, 198, 227
relation to body-weight, 184, 248
Nitrogenous equilibrium, 78
Nitrogenous metabolism, theory of Folin, 144
theory of Pflüger, 138
theory of Voit, 134
Normal diets, 155
influence of chemical character of proteid on, 256
inorganic salts, as aids in, 2
physiological economy in, 264
purpose of, 2
Nutritive balance, as affected by various factors, 117, 118
Nuclease, 71
Nucleoproteid, character of, 3
cleavage products of, 71
Obesity, relation to food consumption, 270
Old age, metabolism in, 296
Osborne and Clapp, on chemistry of proteids of wheat kernel, 258
Osmosis, as factor in absorption, 41
Overeating, evil effects of, 270
Overfeeding, in laying on of fat, 98, 99
Oxidase, xanthin, 73
Oxidases, 64
Oxidation, in metabolism, 60
nature of, in the body, 60
older views regarding, 52
relation to enzymes, 75
site of, in the body, 62
value of respiratory quotient in determination of substances undergoing, 125
views of Lavoisier on, 56
views of Liebig on, 57, 120315
Oxidative cleavage, 61
Oxygen, in katabolism, 62
relation to decompositions in the body, 61
relation to proteid decomposition, 59
Oxygen consumption, in climbing, 116
in relation to work, 123
in standing at rest, 116
in walking, 116
Pancreatic digestion, of proteids, 34
products of, of starch, 37
Pancreatic juice, composition of, 32
condition of trypsin in, 33
enzymes in, 32
sodium carbonate in, 32
Paths of absorption, 44
Pawlow, on adaptation of saliva, 18
Pepsin, in gastric juice, 25, 26
Peptones, 67
absorption of, 41
cleavage by erepsin, 34
effects when injected into blood, 41
formed in gastric digestion, 26
Pflüger, E., theory of proteid metabolism, 138
views on muscle work, 123
Phenol, 37
Phloridzin diabetes, 130
Phosphorus, excretion of, in relation to work, 123
Photograph, of athlete, 190
of Fritz, 199
Photographs, of dogs, 248
of soldiers, 193
Physical endurance, see Endurance
Physiological economy in nutrition, 264
Plastic foodstuffs, 58
Poisons, relation of body resistance to, on low proteid diet, 226
Polypeptid, 35
Portal vein, absorption of food products by, 45
Processes of metabolism, 51
Products, of cleavage of wheat kernel proteids, 259
of gastric digestion, 26
of pancreatic digestion, 37, 67
of proteid cleavage, 70
of putrefaction in intestine, 38
of salivary digestion, 21316
Products of digestion, absorption of, 44
Professional men, fuel value of food on low proteid diet, 178, 180, 185
nitrogen equilibrium of, on low proteid diet, 176, 177, 181
true proteid requirement of, 174
Progeny, influence of meat diet on, in rats, 240
Prosperity, relation to food consumption, 160
Proteid, absorption of, in dogs on low proteid diet, 233, 262
absorption of cleavage products, 47
amounts of food required to supply needs of body for, 272
as food, 3
as fuel, 6
as glycogen former, 130
as source of energy, 122, 123, 124, 129
as source of energy, in fasting, 81
availability of, 12
body-weight on diet low in, 170–175, 181, 185, 190, 199, 245, 249, 250, 251
carbon moiety of, 129
chemical basis of protoplasm, 51
circulating, 134
cleavage products of, 70
consumption by fruitarians, 217
consumption by Japanese, 219, 221
decomposition by oxygen, 59
decomposition in work, 58
excessive consumption of, relation to disease, 269
effect of diet exclusively of, on rats, 239
effect on dogs of diet low in, 233, 234, 237, 245–255
fat absorption in dogs on diet low in, 261
food, real need of body for, 272
formation of carbohydrate from, 129
fuel value of, 15
fuel value of, metabolized during fasting, 86
influence of chemical character of, on nutrition, 256
diet exclusively of, upon progeny of rats, 240
diet low in, on high proteid animals, 231, 233, 243
on excretion of nitrogen, 59, 87, 90
on metabolism, 83
on metabolism of, 88
in foodstuffs, 7
katabolism, 75
luxus consumption of, 59
metabolized during fasting, 86
minimum requirement, 59
morphotic, 134
need of body for, 268
nitrogen equilibrium on diet low in, 176, 177, 181, 200, 201, 245, 249, 250, 251, 252, 255
overfeeding with, 98317
reconstruction of, during absorption, 42
relation of endurance to diet low in, 210, 212
relation of fatigue to diet low in, 208
respiratory quotient of, 107
resistance of body to poisons on diet low in, 226
safety in relation to diet low in, 231
significance of complete cleavage of, 35
strength tests on diet low in, 203, 206
utilization of fat in dogs on diet low in, 261
utilization of nitrogen in dogs on diet low in, 262
work done at expense of, 58
Proteid diet, experiments of Neumann on low, 286
body-weight of dogs on low, 245, 249, 250, 251
body-weight of men on low, 170–175, 181, 185, 190, 199
in relation to nitrogen excretion during hard work, 123, 124
vegetable foods in relation to, 291
Proteid metabolism, influence of carbohydrate on, 92, 94, 95, 96, 97
influence of fat on, 92, 93, 96, 97
influence of proteid on, 59, 87, 90
Folin’s theory of, 144
Pflüger’s theory of, 138
significance of, 131
Voit’s theory of, 134
Proteid requirement, fuel value of food in experiments on, athletes, 188
fuel value of food in experiments on, professional men, 178, 180, 185
fuel value of food in experiments on, soldiers, 198
in dogs, experiments of Jägerroos, 236
in dogs, experiments of Munk, 232
in dogs, experiments of Rosenheim, 234
in dogs, experiments of author, 243
in man, 169, 170, 171, 172, 174–202
nitrogen excretion in experiments on, athletes, 186, 187, 188
nitrogen excretion in experiments on, in dogs, 245, 249, 250, 251, 252, 255
nitrogen excretion in experiments on, professional men, 177, 180, 185
nitrogen excretion in experiments on, soldiers, 197, 200, 201
relation to body-weight, 184, 188, 198, 227
sample diets in experiments on, 178, 182, 189, 195
Proteids, as tissue formers, 58
of wheat kernel, cleavage products of, 259
absorption of, 41
cleavage by erepsin, 34
effects when injected into blood, 41318
Protoplasm, 51
Ptyalin, 20
relation to uric acid, 73
Putrefaction, in intestine, 37
products of, 38
Rats, effects of exclusive proteid diet on, 239
effects of rice on, 240
influence of meat diet on progeny of, 240
Renal activity, and diet, 297
and inorganic salts, 298, 299, 300
Rennin, in gastric juice, 26
Resistance of body to poisons, relation to low proteid diet, 226
Respiration calorimeter, 102
Respiratory foods, 58
Respiratory quotient, 107
influence of foods on, 107, 126
influence of metabolic change on, 108
of foodstuffs, 107
relation to work, 125
value of, in determination of substances oxidized, 125
Rest, carbon dioxide output during, 111
influence of, on oxygen consumption, 126
influence of, on respiratory quotient, 126
Rice, influence of, on growth in rats, 240
Rosenheim, Theodor, on proteid requirement in the dog, 234
Safety of low proteid standards, 231
function of, 20
psychical secretion of, 18
Salivary digestion, in stomach, 23
products of, 21
Salts, see Inorganic salts
Saponification of fats, 36
Sarcosin, 74
Schnyder, 115
Scientific research and typhoid fever, 267
Seasons of the year, relation to diet, 296319
Secretin, 32
Secretion, of gastric juice, 24
Sivén, on proteid requirement, 89
Skatol, 38
Sleep, heat production during, 104, 105
Soaps, 36
Sodium carbonate, in pancreatic juice, 32
Soldiers, fuel value of food in experiments on proteid requirement of, 198
nitrogen equilibrium in experiments on proteid requirement of, 200, 201
photographs of, 193
proteid requirement of, 192
sample diet in experiments on proteid requirement of, 195
strength tests in experiments on proteid requirement of, 203
Specificity of body fat, 44
Standard diets, 155
Standing at rest, oxygen consumption in, 116
Starch digestion, products of, 21, 37
Steapsin, 36
Stomach, absorption from the, 31
as a reservoir, 31
digestion in the, 25–31
fat digestion in the, 36
muscular movements of the, 27–30
salivary digestion in the, 23
time foods remain in the, 29, 30
Storing of proteid, 92, 98, 99, 100
Strength tests, on low proteid diet, athletes, 206
on low proteid diet, soldiers, 203
Sugar, in blood, 45
in blood, relation to glycogen, 46
Sugars, behavior when introduced into blood, 39
selective action in absorption of, 47
Sulphur, excretion of, relation to work, 123
Synthesis, of fat, 43
Temperance in diet, 166, 168, 270
Tissue formers, 58
Tissue metabolism, see Endogenous metabolism
Trypsin, 32
condition in pancreatic juice, 33
Tryptophan, 67
Typhoid fever and scientific research, 267
Underfeeding, dangers of, 214
Urea, 74
excretion of, influence of diet on, 144
relation of, to creatin and creatinin, 74
Uric acid, 73
excretion of, as influenced by diet, 144
relation of, to xanthin bases, 73
Urine, relation of diet to nitrogen distribution in the, 144
Utilization, of dextrose, 40
of disaccharides, 40
of fat in dogs on low proteid diet, 261
of nitrogen in dogs on low proteid diet, 262
Vegetable diet, influence upon dogs, 254, 256
Vegetable foods, relation to low proteid dietary, 291
Vegetables, cellulose in, influence on digestion, 263
Voit, Carl, on minimum proteid requirement, 171
theory of proteid metabolism, 59, 134
Walking, oxygen consumption in, 116
Water in foodstuffs, 7
Watson and Hunter, influence of diet on growth in rats, 239
Wheat kernel proteids, cleavage products of, 259
Weight, see Body-weight
Wislicenus, see Fick and Wislicenus
Work, carbon dioxide excretion in relation to, 123
carbon dioxide excretion during, 111, 112
due to proteid decomposition, 58
effect of, on energy exchange, 109, 110, 113, 115
experiments on man, 110, 111, 112, 113, 114, 115, 116
heat production in, 110
influence of, on oxygen consumption, 126
influence of, on respiratory quotient, 126
nitrogen excretion during excessive, 127
nitrogen excretion during fasting in, 125
proteid decomposition in, 58
relation of diet to, 126
to energy exchange, 119
fats and carbohydrates to, 58
nitrogen excretion on proteid diet to hard, 123, 124
nitrogen excretion to proteid diet to hard, 122, 123, 124321
relation of oxygen consumption to, 123
phosphorus excretion to, 123
sulphur excretion to, 123
respiratory quotient in relation to, 125
source of energy during fasting in, 125
views of Argutinsky on muscle, 123
views of Pflüger on muscle, 123
views of Voit on muscle, 59, 134
Xanthin, 72
Xanthin oxidase, 73
1 Edward Curtis, M.D. Nature and Health: Henry Holt & Co., New York. 1906. p. 39.
2 The data composing this table are taken from Bulletin 28 (Revised Edition), United States Department of Agriculture, Office of Experiment Stations.
3 Stohmann: Ueber den Wärmewerth der Bestandtheile der Nahrungsmittel. Zeitschr. f. Biol., Band 31, p. 373.
4 See Rubner: Calorimetrische Untersuchungen. Zeitschr. f. Biol., Band 21, p. 250. Also, Rubner: Die Quelle der thierischen Wärme. Ibid., Band 30, p. 73.
5 Pawlow: The Work of the Digestive Glands. Translated by Thompson. London, 1902.
6 Emil Fischer: Bedeutung der Stereochemie für die Physiologie. Zeitschr. für physiologische Chemie, Band 26, p. 60.
7 The Physiology of Digestion. By William Beaumont, M.D. Second Edition, 1847, p. 100.
8 W. B. Cannon: The Movements of the Stomach studied by means of the Röntgen Rays. American Journal of Physiology, vol. 1, p. 359.
9 W. B. Cannon: The Passage of different Food-stuffs from the Stomach and through the Small Intestine. American Journal of Physiology, vol. 12, p. 387.
10 Emil Abderhalden: Abbau und Aufbau der Eiweisskörper im thierischen Organismus. Zeitschr. f. physiologische Chemie, Band 44, p. 27.
11 Otto Cohnheim: Zur Spaltung des Nahrungseiweisses im Darm. Zeitschrift f. physiologische Chemie, Band 49, p. 64.
12 Bergell and Lewin: Zeitschrift für experimentelle Pathologie und Therapie, Band 3, p. 425.
13 Franz Hofmeister: Ueber Resorption und Assimilation der Nährstoffe. Archiv f. d. exper. Pathol. u. Pharm., Band 25, p. 240.
14 J. B. Leathes: Problems in Animal Metabolism. Blakiston’s Son and Co., 1906, p. 123.
15 Taken from Sir Michael Foster’s “Lectures on the History of Physiology during the Sixteenth, Seventeenth, and Eighteenth Centuries.” Cambridge, 1901, p. 12.
16 See Foster’s Lectures, p. 136.
17 Quoted from Foster’s Lectures, p. 195.
18 See C. Voit: Hermann’s Handbuch der physiologie des Gesammt-Stoffwechsels. Band 6, Theil 1, p. 269, 1881.
19 Loc. cit., p. 270.
20 See M. Jacoby: Ueber die Bedeutung der intracellulären Fermente für die Physiologie und Pathologie. Ergebnisse der Physiologie, Erster Jahrgang, 1. Abtheilung, p. 230.
21 These data were furnished the writer by Dr. Thomas B Osborne, and represent in large measure the results of his own chemical work.
22 Taken from Landergren: Untersuchungen über die Eiweissumsetzung des Menschen. Skandinavisches Archiv für Physiologie, Band 14, p. 112; and from A Magnus-Levy: v. Noorden’s Handbuch der Pathologie des Stoffwechsels, 1906, p. 312.
23 Expressed in this form from Voit’s figures by A Magnus-Levy. Loc. cit., p. 311.
24 Taken from Johansson, Landergren, Sondén, and Tiegerstedt: Beiträge zur Kenntniss des Stoffwechsels beim hungernden Menschen. Skandinavisches Archiv für Physiologie, Band 7, p. 29.
25 The carbon output represents the total carbon of the expired air, urine, and excrement.
26 Leathes: Problems in Animal Metabolism. Philadelphia, 1906, p. 157.
27 Sivén: Zur Kenntniss des Stoffwechsels beim erwachsenen Menschen, mit besonderer Berücksichtigung des Eiweissbedarfs. Skandinavisches Archiv für Physiologie, Band 11, p. 308.
28 C. Voit: Hermann’s Handbuch der Physiologie des Gesammtstoffwechsels, Band 6, p. 130.
29 Landergren: Untersuchungen über die Eiweissumsetzung des Menschen, Skandinavisches Archiv für Physiologie, Band 14, p. 114.
30 An experiment by Miura, quoted from A. Magnus-Levy in v. Noorden’s Handbuch der Pathologie des Stoffwechsels, 1906, p. 331.
31 Max Dapper: Ueber Fleischmast beim Menschen. Inaug. Disser. Marburg, 1902.
32 Zeitschrift für klinische Medizin, Band 44, p. 22.
33 For an account of the respiration calorimeter and the great diversity of apparatus accessory thereto, together with a description of the methods of measurement, analysis, etc., see Publication No. 42, Carnegie Institution of Washington, “A Respiration Calorimeter with Appliances for the Direct Determination of Oxygen.” By W. O. Atwater and F. G. Benedict.
34 In the experiment, the body lost 29.16 grams of proteid = 165 calories, but gained fat and glycogen = 393 calories. Hence, there were 229 calories gained from body material.
35 See Armsby: Principles of Animal Nutrition, p. 368.
36 Taken from Armsby: Principles of Animal Nutrition, p. 383.
37 Atwater and Benedict: Experiments on the Metabolism of Matter and Energy in the Human Body 1900–1902. Bulletin No. 136, Office of Experiment Stations, U. S. Department of Agriculture, 1903, p. 141.
38 Loc. cit., pp. 130 and 131.
39 See W. O. Atwater and H. C. Sherman: The effect of severe and prolonged muscular work on food consumption, digestion, and metabolism. Bulletin No. 98, Office of Experiment Stations, U. S. Department of Agriculture.
40 Atwater and Sherman. Loc. cit., p. 51.
41 L. Schnyder: Muskelkraft und Gaswechsel. Zeitschrift für Biologie, Band 33, p. 289.
42 G. Katzenstein: Ueber die Einwirkung der Muskelthätigkeit auf den Stoffverbrauch des Menschen. Pflüger’s Archiv für die gesammte Physiologie, Band 49, p. 330. Also Magnus-Levy: v. Noorden’s Handbuch der Pathologie der Stoffwechsel, p. 233.
43 See Gesammelte Schriften von Adolf Fick. Ueber die Entstehung der Muskelkraft. Band 2, p. 85. Würzburg, 1903.
44 Pflüger: Die Quelle der Muskelkraft. Pflüger’s Archiv für die gesammte Physiologie, Band 50, p. 98.
45 Argutinsky: Muskelarbeit und Stickstoffumsatz. Ibid., Band 46, p. 552.
46 Bunge: Textbook of Physiological and Pathological Chemistry. Second English Edition, 1902, p. 352.
47 Quoted from Leathes: Problems in Animal Metabolism, p. 100.
48 This and the following account of Weston are taken from Bulletin No. 98, U. S. Department of Agriculture, Office of Experiment Stations. The effect of severe and prolonged muscular work on food consumption, digestion, and metabolism. By W. O. Atwater and H. C. Sherman, p. 13.
49 See Leo Langstein: Die Kohlehydratbildung aus Eiweiss. Ergebnisse der Physiologie, Band 3, Erster Theil, p. 456.
See also, Lüthje: Zur Frage der Zuckerbildung aus Eiweiss. Archiv für d. gesammte Physiologie, Band 106, p. 160.
50 See Voit: Hermann’s Handbuch der Physiologie, Band 6, p. 301.
51 Eduard Pflüger: Ueber einige Gesetze des Eiweissstoffwechsels (mit besonderer Berücksichtigung der Lehre vom sogenannten “circulirenden Eiweiss”). Archiv f. d. gesammte Physiologie, Band 54, p. 333.
52 Otto Folin: Laws Governing the Chemical Composition of Urine. American Journal of Physiology, vol. 13, p. 66. A theory of Protein Metabolism. Ibid., vol. 13, p. 117.
53 Bulletin No. 149. Woods and Mansfield. Studies of the Food of Maine Lumbermen. U. S. Department of Agriculture, 1904.
54 Bulletin No. 75. Atwater and Bryant. Office of Experiment Stations, U. S. Department of Agriculture, 1900.
55 Felix Hirschfeld: Untersuchungen über den Eiweissbedarf des Menschen. Pflüger’s Archiv f. d. gesammte Physiologie, Band 41, p. 533.
56 Carl Voit: Ueber die Kost eines Vegetariers. Zeitschrift für Biologie, Band 25, p. 232.
57 Klemperer: Untersuchungen über Stoffwechsel und Ernährung in Krankheiten. Zeitschrift für klin. Medizin, Band 16, p. 550.
58 L. Breisacher: Ueber die Grösse des Eiweissbedarfs beim Menschen. Deutsche med. Wochenschrift. 1891. No. 48.
59 W. Caspari: Physiologische Studien über Vegetarianismus. Bonn. 1905. p. 13.
60 In presenting the general results of these experiments, the writer has drawn freely from his book, “Physiological Economy in Nutrition,” published by the Frederick A. Stokes Company, New York, 1904.
61 In presenting the general results of these experiments, the writer has drawn freely from his book, “Physiological Economy in Nutrition,” published by the Frederick A. Stokes Company, New York, 1904.
62 The coffee was prepared with milk and sugar.
63 Through the kindness of Professor Fisher, the writer has had the opportunity of reading the report of this work, which at this writing is not published, and he has drawn upon it freely for the following statements of fact.
64 Edward Curtis, M. D.: Nature and Health. New York, Henry Holt & Co. 1906. p. 71.
65 Bulletin No. 107, Office of Experiment Stations, U. S. Department of Agriculture, 1901, from which the descriptions given have been taken.
66 A Digest of Japanese Investigations on the Nutrition of Man. Bulletin No. 159, Office of Experiment Stations, U. S. Department of Agriculture, 1905.
67 Ueber die Folgen einer ausreichenden, aber eiweissarmen Nahrung. Ein Beitrag zur Lehre vom Eiweissbedarf. Virchow’s Archiv für pathologische Anatomie und Physiologie, Band 132, p. 91.
68 Theodor Rosenheim: Ueber den Gesundheitsschädigenden Einfluss eiweissarmer Nahrung. DuBois-Reymond’s Archiv für Physiologie, 1891, p. 341. Also, Weiterer Untersuchungen über die Schädlichkeit eiweissarmer Nahrung. Pflüger’s Archiv f. d. gesammte Physiologie, Band 54, p. 61, 1893.
69 B. H. Jägerroos: Ueber die Folgen einer ausreichenden, aber eiweissarmen Nahrung. Skandinavisches Archiv für Physiologie, Band 13, p. 375, 1902.
70 Chalmers Watson, M.D., and Andrew Hunter, M.B.: Observations on Diet. The Influence of Diet on Growth and Nutrition. Journal of Physiology, Vol. XXXIV, p. 112, 1906.
71 The fuel value of the food was calculated from the data given in Bulletin No. 28, U. S. Department of Agriculture. All figures for nitrogen were obtained by exact chemical analysis.
72 All through the balance periods the dogs were catheterized each morning to insure complete collection of the twenty-four hours’ urine.
73 See Osborne and Clapp: The Chemistry of the Protein Bodies of the Wheat Kernel. American Journal of Physiology, vol. 17, p. 231.
74 There is an unavoidable error here, since the excrement contains not only undigested food, but also contains some nitrogenous matter derived from the secretions of the intestine, etc.
75 Edward Curtis, M.D.: Nature and Health, p. 70. Henry Holt & Company, New York, 1906.
76 Aran Coirce: British Medical Journal, April 7, 1906, p. 829.
77 Irving Fisher: A new method for indicating food values. American Journal of Physiology, vol. 15, p. 417, 1906.
78 Aran Coirce: British Medical Journal, April 7, 1906, p. 829.
79 Dr. med. et phil. R. O. Neumann: Experimentelle Beiträge zur Lehre von dem täglichen Nahrungsbedarf des Menschen unter besonderer Berücksichtigung der notwendigen Eiweissmenge. Archiv für Hygiene, Band 45, p. 1, 1902.
80 C. A. Herter: Character of the Bacterial Flora of Carnivorous and Herbivorous Animals. Science, December 28, 1906, p. 859.
81 Dr. S. Fenger: Beiträge zur Kenntniss des Stoffwechsels im Greisenalter. Skandinavisches Archiv für Physiologie, Band 16, p. 222, 1904.