
Title: Piebald rats and selection
An experimental test of the effectiveness of selection and of the theory of gametic purity in Mendelian crosses
Author: William E. Castle
John C. Phillips
Release date: September 5, 2024 [eBook #74375]
Language: English
Original publication: Washington, DC: Carnegie Institution of Washington, 1914
Credits: Brian Wilson, Bryan Ness and the Online Distributed Proofreading Team at https://www.pgdp.net (This file was produced from images generously made available by The Internet Archive/Canadian Libraries)
PIEBALD RATS AND SELECTION
AN EXPERIMENTAL TEST OF THE EFFECTIVENESS OF
SELECTION AND OF THE THEORY OF GAMETIC
PURITY IN MENDELIAN CROSSES
BY
W. E. CASTLE and JOHN C. PHILLIPS
WASHINGTON, D. C.
Published by the Carnegie Institution of Washington
1914
Carnegie Institution of Washington, Publication No. 195
Paper No. 21 of the Station for Experimental Evolution
at Cold Spring Harbor, New York
From the Laboratory of Genetics
of the Bussey Institution
Copies of this Book
were first issued
FEB 18 1914
PRESS OF GIBSON BROTHERS, INC.
WASHINGTON, D. C.
| Page. | |
| Introduction | 5 |
| Material and methods | 7 |
| Plus selection series | 9 |
| Minus selection series | 12 |
| Return selection | 13 |
| Crosses with wild rats | 16 |
| Crosses with black “Irish” rats | 18 |
| Plus selection of “extracted hooded” rats | 20 |
| Crosses of the plus race with the minus race | 22 |
| Summary of results | 22 |
| Discussion | 23 |
| The “mutant” series | 25 |
| Bibliography | 31 |
| Tables | 32-54 |
| Explanation of plates | 56 |
The fundamental importance of Mendel’s law of heredity is generally recognized among biologists. It is a working hypothesis whose utility is fully substantiated by abundant results daily increasing in amount. But biologists are not in agreement as to how much this law includes. All perhaps would agree that it implies the existence in the germ-cell of specific determiners essential for the production of particular characteristics in the offspring. Further, no one probably will object to the statement that it implies a dual or duplex condition of the zygote as regards determiners and a simple or simplex condition of the gamete. Thirdly, the fact will be admitted by all that most mendelizing characters are wholly independent of each other in heredity, for which reason we are forced to suppose that their determiners are distinct within the germ-cell.
But beyond these few generalizations great diversity of opinion exists. As regards the very nature and function of the determiners, some consider them unvarying, and explain the observed variation of mendelizing characters in organisms as due to a modifying action of other determiners. At one time even a modifying action of other determiners was denied, and the theory was advanced that the gametes extracted from a mendelian cross are pure as regards the single characters which may have been concerned in that cross. Investigations carried out by Castle have done something to dispel this idea. In particular it was shown (Castle, 1905, 1906; Castle and Forbes, 1906) that in guinea-pigs, polydactylism, long-hair, and rough coat are mendelizing characters which are affected in the degree of their development by crosses—that is, when these characters are “extracted” from crosses the characters are not exactly the same as before; hence the gametes are not “pure.”
The experimental result is not denied, but in order to save the substance of the theory its advocates now suppose that the determiners have not changed, but in consequence of the cross certain modifiers have become associated with them which change their appearance in the organism. The real unchanging thing is now called the “genotype,” its appearance the “phenotype.”
In this genotype theory we are dealing only with a new and more refined aspect of the “theory of pure gametes.” It is not a necessary part of mendelism, not even an original part; but it is very important for us to know whether it is true or not. For if it is true, selection unattended by hybridization is largely a waste of time, as De Vries and Johannsen have maintained, and Jennings and Pearl have reiterated.
The investigation which we are about to describe was started six years ago to test the validity of the theory of pure gametes which was then current. Pure “genes” had not yet been invented. The investigation has been in continuous progress ever since, and while we expect to continue it further, it seems to us desirable that the results already obtained be presented for criticism.
Some conception of the work entailed in the investigation may be gathered from the statement that we have during its progress reared and studied the color pattern of over 25,000 rats. A long and arduous investigation of this kind has been made possible by a series of grants from the Carnegie Institution of Washington made to the senior author, for which he here makes grateful acknowledgment. Thanks are also due to Dean W. C. Sabine, of Harvard University, for encouraging and supporting the work in a variety of ways.
In June 1906 Dr. Hansford MacCurdy completed, under the direction of the senior author, a study of the inheritance of color in rats. His studies had shown that the piebald pattern of “hooded” rats behaves as a mendelian recessive character in relation to the uniform or nearly uniform coloration of wild rats, but that the hooded pattern, when extracted from a cross with wild stock, shows a different variability, the pigmentation of the extracted recessives being increased in extent. This result was interpreted as showing the unsoundness of the current doctrine of “purity of the gametes” in mendelian crosses.
Upon the conclusion of Dr. MacCurdy’s experiments, the pedigreed stock which he had used was not entirely discarded. A certain portion of it was utilized for new experiments designed to show whether the “hooded” coat-pattern can be modified by selection unattended by cross-breeding.
Two series of selections were started in October 1907, in one of which animals were chosen as parents which had pigmentation as extensive as possible. This we may call the plus series. In the other series animals were chosen as parents which had pigmentation as restricted as possible. This we may call the minus series.
During the academic year 1906-7, the experiments were in immediate charge of Mr. W. G. Vinal; during 1907-8 the plus series was in charge of Mr. H. S. Rand, while the minus series was in charge of Mr. F. C. Bradford. Throughout this time the experiments were closely supervised by the senior author, who assisted in the “grading” of every litter of young. In October 1908 the junior author began his association in the experiments, which has continued up to the present time. Throughout these five years he has looked after the details of the experiments almost continuously, but both authors have in most cases taken part together in the grading of the young, and in no case has the grading been done except under the immediate supervision of one or the other of the authors. This fact is stated to show that the personal element in the grading has been kept as constant as possible. In the tabulation of results and computation of statistical constants, the authors have worked together. This statement of results is written by the senior author.
During the year 1906-7 the young rats were graded by the method used by MacCurdy and Castle (1907) that is, the back-stripe was measured and a calculation made of the percentage of the dorsal surface posterior to the hood which was pigmented. But on account of the irregular outline of the back-stripe in many individuals the method of measurement was found to be at best a rough one, as well as extremely laborious. Accordingly in the summer of 1907 a set of arbitrary grades[8] was adopted, which is shown at the top of Plate 1. Each young rat was classed in that grade which it most nearly approached in amount of pigmentation. Skins of rats graded from -3¼ to +4¾ are shown in the middle and lower rows of Plate 1. The grading was done when the rats were about three or four weeks old, at which time selected individuals were reserved as the parents for a later generation, the remainder being discarded. This method has been followed ever since its adoption and the data thus obtained are summarized in the tables, which cover the breeding operations of a little more than six years, 1907-1913.
The grouping of the young in a series of generations is only approximately accurate, for practical considerations have often led us to mate together animals which belonged to different generations of offspring. When, for example, an animal of generation 2 was mated with one of generation 4, the question would arise: To what generation do the offspring belong? In deciding this question we simply added one to the mean of the generations to which the respective parents belonged. In the foregoing case this would be (2 + 4)/2 + 1 = 4.
In case one parent belonged to generation 2 and the other to generation 3, a fractional result would be obtained, thus (2 + 3)/2 + 1 = 3½. In making up the summaries of the generations as given in the tables, offspring like the foregoing, of generation 3½, were divided equally between generations 3 and 4, alternate litters of young as recorded in the ledger being assigned to each. Offspring belonging to generations 2¾ and 3¼ were tabulated in generation 3; those belonging to generations 3¾ and 4¼ were tabulated in generation 4, etc. While, therefore, the generations as tabulated overlap, it is clear that they include groups of offspring of selected parents each the result of one additional selection over the preceding group.
The early generations include too few individuals to be of much statistical value, but where the number of offspring rises to 500 or over, the statistical constants acquire undoubted value. The data have been given in the form of correlation tables which will repay careful study. In the tables a single entry has been made for each individual offspring in that row which corresponds with the mean grade of its two parents. Thus, if one parent were of grade 2 and the other of grade 2½, the offspring would be entered in the row 2¼ along with the offspring of parents both of grade 2¼. Offspring of parents whose mean grade fell between the rows given in the tables were divided equally between the adjacent rows, alternate litters being assigned to each. Thus, if the mean grade of the parents were 2⅟₁₆, alternate litters of offspring would be entered in row 2 and in row 2⅛.
This series begins with pairs ranging in average grade from +1.87 to +3. From these parents were obtained 150 young, which range in grade from +1 to +3, as is shown in Table 1. It will be observed that the lower-grade parents have on the average lower-grade offspring than the higher-grade parents. But in no case is the average grade of the offspring as great as that of their parents. Thus 1.87 parents had 1.82 offspring (average grade); 2.00 parents had 1.76 offspring; 2.25 parents had 1.87 offspring; and so on to 3.00 parents, which had 2.35 offspring. There is a falling back in grade or “regression” of the offspring as compared with their parents, which increases in amount as the grade of the parents becomes higher. (See column “Regression” in Table 1.) The parents of this first generation were chosen because of their high grade. They were all probably in grade above the general average of the population from which they were selected. In the case of those which deviate most from the general average the regression is greatest, as we should expect.
This phenomenon of regression, which is a very general one in cases of selection, was first observed by Galton in selecting sweet-peas of varying size from a mixed population. Later Johannsen, who repeated the experiment with beans, found that by pedigree culture he was able to break the mixed population up into pure lines within which, considered singly, no regression occurred. We shall need later to return to this subject and consider whether pure lines free from regression exist or can be produced as regards the hooded pattern of rats.
Returning to the examination of Table 1, since the high-grade parents produce higher-grade offspring than do the low-grade parents, it is evident that we might hope by further selection either to isolate a pure line of high-grade rats which would be free from regression and therefore stable, or else to advance the grade of the offspring still higher, even though regression persists. As a measure of the extent to which high-grade parents have high-grade offspring and vice versa, in each generation, we may employ the well-known correlation coefficient. This for Table 1 is 0.30.
The second generation in the plus series (Table 2) includes the offspring of parents which appear as offspring of the higher grades in Table 1, together with a few individuals which appear in Table 2 both as offspring and as parents of other offspring, by reason of their having been mated with generation 1 individuals and so having produced generation 1½ offspring, as explained on page 8. To obtain larger numbers of offspring, several new pairs were added to the experiment in this generation, which do not appear in Table 1 either as offspring or as parents, but which were derived from the same general stock as the parents of generation 1. Their inclusion here accounts for the very[10] low range of the offspring in Table 2, which extends from -1.00 to +3.75. The parents’ range (means of pairs) extends from 2.00 to 3.12. The grand average of the parents is 2.52, that of the offspring is 1.92. The correlation between grade of parents and grade of offspring is 0.32.
From this point on in the series no new stock was added and each generation of offspring furnished the parents for the following generation, except for the slight overlapping of generations when parents of different generations were mated with each other, as has already been explained.
In generation 3, Table 3, the parents ranged from 2.12 to 3.37 in grade, the offspring from 0.75 to 4.00. The mean of the parents was 2.73, that of the offspring 2.51. The degree of correlation between parents and offspring is expressed by the coefficient 0.33 (a perfect correlation would give 1.00).
In generation 4, Table 4, the selection of parents became considerably more rigid; most of the parental pairs were of grade 3 or higher, their average being 3.09. The average grade of the offspring was 2.73, their range extending from 0.75 to 3.75. The correlation in this generation fell very low, to 0.07, not because of a lessened regression but rather because of a very high regression on the part of the offspring of high-grade parents.
In generation 5, Table 5, the grade of the selected parents ranged from 2.75 to 4.12, its mean being 3.33. The offspring, showing the usual regression, ranged from 0.75 to 4.25, their mean grade being 2.90. The correlation between parents and offspring in this generation was 0.16. The number of individuals comprising this generation of offspring was 610.
It is scarcely necessary to discuss separately the correlation table for each of the next eight generations, Tables 6 to 13. The number of offspring rises to a maximum (1,408) in generation 8, Table 8; then declines to less than 200 in generation 13. But as this generation and the preceding one are still being produced, it is probable that the number recorded will be considerably increased before the generation is complete. The means of parents and offspring and the other statistical constants for the several generations can be most easily compared by reference to Table 14. Leaving out of consideration the exceptional generation, 2, the following will be observed:
(1) The mean of the selected parents has steadily advanced throughout the series, as has also that of their offspring.
(2) The variability (standard deviation) of the parents as a group has decreased somewhat as increase in numbers made a more rigid selection possible; that of the offspring has undergone a similar change.
(3) The correlation between parents and offspring has not materially changed. The average of the correlation coefficients for the entire series is 0.194, for the last three generations it is 0.175, for the three preceding[11] generations it is 0.141, for the three which precede those it is 0.185, while for the first four generations it is 0.253. In every case the correlation is positive—that is, the higher-grade parents have higher-grade offspring and vice versa.
(4) The offspring as a group average lower in grade than their parents—that is, their mean regresses on that of the selected parents, but because of the higher mode about which variation occurs in each generation certain of the offspring are of higher grade than their parents. Thus an elevation of the grade of the parents in the next generation is made possible.
(5) With the selection of more extreme parents, the absolute regression of the offspring has not increased, but on the contrary has slightly diminished—that is, the advance made by the parents is retained by their offspring.
In Table 15 have been brought together for comparison the means of the several horizontal rows of Tables 1 to 13. By examining the vertical columns of Table 15 the mean grade of the offspring of parents of a particular grade in any generation may be compared at a glance with that of parents of the same grade in any other generation. By running the eye down the columns, it will be observed that the mean grade of the offspring tends to increase upon repeated selection. Thus parents of grade 3.75 appear first in generation 4, the grade of their offspring being 2.75; the offspring of such parents in subsequent generations grade in order, 3.07, 3.22, 3.35, 3.49, 3.50, 3.69, 3.75, and 3.83 (twelfth generation not complete). The difference between parents and offspring in this series grows less and less and finally disappears altogether. If the grade of 3.75 parents in this series is compared with the grade of all offspring in the corresponding generations we have the following:
| Generation. | Mean of offspring of 3.75 parents. | Mean of offspring of all parents. |
|---|---|---|
| 4 | 2.75 | 2.73 |
| 5 | 3.07 | 2.90 |
| 6 | 3.22 | 3.11 |
| 7 | 3.35 | 3.20 |
| 8 | 3.49 | 3.48 |
| 9 | 3.50 | 3.54 |
| 10 | 3.69 | 3.73 |
| 11 | 3.75 | 3.77 |
| 12 | 3.83 (35 individuals) | 3.94 (590 individuals) |
In generation 4 the 3.75 parents represented the most advanced individuals of the series, a whole grade in advance of the general average of the race. Their offspring showed a correspondingly large regression. The general average of the race steadily advanced in later generations until in generation 11 it equaled that of the 3.75 parents; then the regression vanished. In the following generation, 12 (which is still incomplete, but in which the average of the offspring thus far is 3.94), the 3.75 group of parents, which are now below the average of the race,[12] actually produce offspring of higher grade than themselves, viz, 3.83. It will thus be seen that the regression is uniformly toward the mean of the race and changes its direction when that mean changes its position with reference to a particular grade of parents. This conclusion is supported by other columns of Table 15, but is best illustrated by this particular case because here the selection has extended over a greater number of generations than elsewhere in the series.
If one examines the horizontal rows of Table 15, he finds in general that numbers increase toward the right. Exceptions are commonest toward the ends of the rows where fewest individuals are represented. This increase means that, within any generation, as the grade of the parents rises, that of their offspring rises also. Since in general the selected parents are above the general average of the race for the time being, regression is naturally downward in nearly all cases.
From what precedes we may conclude (1) that in this series of rats the somatic character (appearance) of an individual is in general a true indication of its germinal character, since the higher the grade of the parents the higher the grade of the offspring, and vice versa; but that (2) the somatic character of an individual is not a perfect index of its germinal character, since the offspring of aberrant individuals are less aberrant than themselves, i. e., the offspring regress toward the mean of the race; yet that (3) by selection of plus variations we can displace, in a plus direction, not only the mean of the race, but also the upper and lower limits of its variation, the total amount of variability (standard deviation) being thereby only slightly decreased.
This series begins with selected parents ranging in grade from -1.25 to -1.87. Their average, if each pair is weighted in proportion to the number of its offspring, is -1.46. The offspring (Table 16), like the offspring of the original plus selections, regress toward grade 0. They range in grade from +0.25 to -2.00, their mean being -1.00. The total number of offspring recorded in this generation is only 55, this being too small to warrant the calculation of a correlation coefficient.
Generation 2 (Table 17) is somewhat larger, but still too small to make statistical constants based upon it of much consequence. The offspring show substantially the same range of variation as in the previous generation, but with a slightly higher average (-1.07). The coefficient of correlation (-0.03) is negative, but too small to be significant. The record of the next eleven generations will be found summarized in Tables 18 to 28, or in more condensed form in Tables 29 and 30. Generation 13 (Table 28) is still incomplete.
The mean of the parents steadily rises from -1.56 in generation 3 to -2.50 in generation 13. The mean of the offspring rises by like increments from -1.18 in generation 3 to -2.39 in generation 13. There is[13] throughout these generations a positive correlation between parents and offspring. This amounts on the average to 0.137 as compared with 0.193 observed in the plus selection series. The absolute change in amount of pigmentation is no doubt less in the minus selection than in the plus selection series, but if the change were recorded as percentage decrease of pigmentation in one case and percentage increase in the other, the change indicated would probably be as great in one as in the other.
In the minus as in the plus series we observe:
(1) The character of the offspring varies with that of the parents; high-grade parents have high-grade offspring and vice versa.
(2) The variability of the race (as indicated by the standard deviation) undergoes some reduction and the limits of variation, both upper and lower, are displaced in the direction of the selection.
(3) The regression from a new and extreme class of parents is at first large, but decreases as the selection is repeated and finally disappears altogether when the average of the race becomes equal to the particular grade under discussion.
The plus and minus selection series already described make it clear that one can, in a race of hooded rats, either increase or decrease the average pigmentation at will, and at the same time secure more advanced stages either of pigmentation or of depigmentation than those previously occurring in the race. The question now arises, are these changes permanent; will these displaced means retain their new position, if the race is left to itself; or will the newly obtained stages vanish as soon as selection is suspended? A presumption that the changes will prove permanent is afforded by the gradual decrease of regression and its final reversal in the case of offspring of a particular grade, upon repeated selection made in the same direction. (See page 12.) But in order to test the matter more directly and thoroughly, the experiment has been repeatedly made of reversing the course of selection, after it had been in progress for several generations, with a view of ascertaining whether the return toward the former condition of the race would be made more speedily and easily than the original departure from it had been.
The first experiment of this sort was a return selection from generation 6 (and 6½) of the minus selection series. The parents of generation 6 (Table 21) averaged -1.86 in grade; the average grade of their offspring was -1.56, a regression of 0.30. The range of the offspring extended from 0 to -2.50. Some low-grade offspring were chosen for a return selection series (Table 31). The mean grade of the selected pairs ranged from -0.37 to -0.87, their mean being -0.60. These parents produced 118 offspring, whose average grade was -1.28, a regression of 0.68 in a direction contrary to that of the regression in the[14] minus selection series. The large amount of the regression might seem to imply that it was even more difficult to return toward the former state of the race (in the neighborhood of 0) than it had been to depart from it, but this can not be insisted on, because the number of individuals under observation is not sufficiently large. To test the reality and permanency of the reversed regression, the selection was repeated five additional times, altogether six successive return selections being made with the idea of undoing what had been effected by six original selections in an opposite direction. The result of the second successive return selection is shown in Table 32. The parents here were of grade -0.50 and they produced 19 offspring of the average grade -0.95, a regression of 0.45 away from 0 as before.
Table 33 shows the result of the third return selection. Individuals entered in Table 32 as offspring appear here as parents. Only those pairs which were of mean grade, -0.25 or -0.37, should really be regarded as a third return selection. They gave offspring with mean grades of -0.63 and -0.86 respectively, which show regression of 0.38 and 0.49 away from 0.
But Table 33 shows also the character of young produced by -1.12 and -1.25 parents in this same third return-selection generation, i. e., by unselected parents of the generation in question. Their young also regress away from 0—that is, in the direction of the original selection. The -1.12 parents produced -1.61 offspring, a regression of 0.49, while the -1.25 parents produced -1.35 offspring, a regression of 0.10. For Table 33 as a whole the regression away from 0 averages 0.31.
A fourth generation in the return-selection series is summarized in Table 34. The parents are of mean grade -0.63; their 50 offspring are of mean grade -1.17, a regression amounting to 0.54 away from 0 and in the direction of the six generations of original selection.
Table 35 contains the results of the fifth generation of the series. The parents are here of mean grade -0.65. The number of offspring is very small (13), but they nevertheless show the reversed regression which characterized the four preceding generations. Their mean was -0.75, a regression of 0.10 away from 0.
A sixth and final generation in this return-selection experiment is summarized in Table 36. It includes 36 offspring of mean grade -0.39, the mean of the parents being -0.26, a regression of 0.13 away from 0. It will be seen, therefore, that the effect of the six original selections had not been entirely overcome by an equal number of return selections. The reason for this is obvious. Much smaller numbers are concerned in the return selections than in the original minus selections. The return selections are accordingly less efficient. Nevertheless, after the sixth return selection we find that 1 in 6 of the offspring have plus grades and their average is lower (that is, less minus) than the offspring in the minus series after a single generation of selection. (Cf. Tables 16 and 36.)
The amount and persistency of the reversed regression in this series show clearly that return selection is not easier or more rapid than the original modification of the race by selection, but that selection in either a plus or minus direction has cumulative and permanent effects.
Further support for this conclusion is furnished by return selections (one each) made from the seventh generation, from the eighth generation, and from the eleventh generation of the minus selection series. (See Tables 37, 38, and 39.) Generation 7 (Table 22) was produced by parents of average grade -2.01. Their offspring were of average grade -1.73, a regression (toward 0) amounting to 0.28. Certain pairs of these offspring of grade -0.75 and -0.87 (mean -0.78) constitute the return selection from generation 7 (Table 37). They had 33 offspring of average grade -1.15, a regression away from 0 amounting to 0.37.
Generation 8 of the minus-selection series (Table 23) was produced by parents of mean grade -2.05. Their offspring were of mean grade -1.80, a regression (toward 0) of 0.25. Certain pairs of these offspring of grades -0.50, -0.62, and -1.00 (mean -0.72), when chosen as parents, produced 41 young of mean grade -1.51, a regression away from 0 amounting to 0.79. (See Table 38.)
Generation 11 of the minus series (Table 26) was produced by parents of mean grade -2.30. The offspring were of mean grade -2.15, a regression of 0.15 toward 0. A pair of the offspring of mean grade -1.62 (Table 39) produced 16 young of mean grade -1.95, a regression of 0.32 away from 0. This result shows that the selected race had now passed the point represented by the grade of the parents (-1.62) and the offspring regressed toward a racial mean as advanced as the most extreme individuals obtained previous to selection.
To show that, in the plus selection series, a return selection has a result similar to that just described, two experiments may be cited:
The sixth generation of the plus selection series was produced by parents of mean grade 3.52, and their offspring were of mean grade 3.11, a regression toward 0 amounting to 0.41. Certain of these offspring of mean grade 2.00, when chosen as parents, produced 17 young of mean grade 2.36, a regression away from 0 amounting to 0.36. (See Table 40.)
The eleventh generation of the plus selection series (Table 11) was produced by parents of mean grade -3.97; their offspring were of mean grade -3.78, a regression of 0.19 toward 0. Certain of these offspring, ranging in grade from -2.62 to -3.25 (Table 41), mean -2.79, produced 53 young of mean grade -3.32, a regression away from 0 amounting to 0.53. The regression in this case, as in all those previously described, was toward the racial mean of the previous generation, which, however, it has in no case reached.
This can have but one meaning. The genetic character of the hooded rat is in a general way correctly indicated by its somatic character. Selection is therefore immediately effective, whether plus or minus in character, and whether or not preceded by selection in the same direction or in an opposite direction. But regression may be expected from the character of aberrant parents back toward the normal of the previous generation, yet this regression will in general be less than the departure of the aberrant parents from the normal of their generation. If one desires in such a case to obtain continuous and progressive departure from the normal in either a plus or a minus direction, he need only select continuously in the desired direction.
As a further test of the permanency of the modification effected by selection in the hooded pattern of rats, crosses have from time to time been made of the selected races with a pure wild stock, i. e., with ordinary wild animals caught in traps. In all cases the wild animals used were known to be homozygous as regards gray coat and self pattern, since when crossed with black-hooded animals they produced only gray self offspring. In such crosses the hooded pattern is recessive, the F₁ offspring being indistinguishable from ordinary wild gray rats except for the possession of a white patch of varying size upon the belly, but even this may be lacking. (See Plate 2, ♂ 8000, 8018, and 8021.)
The grade of the hooded young extracted from a cross with wild animals corresponds in a general way with the grade of the hooded animal used in making the cross, as the following cases will show. (Compare also Plates 2 and 3.)
A female of grade -1.87, belonging to generation 2½, minus series (compare Tables 2 and 3), was crossed with a wild male. (See Plate 2, ♀ 6176.) Among her F₂ descendants (cf. Plate 2, 8070 to 8078) occurred 62 hooded individuals, whose grade distribution is shown in Table 42, first row. Their mean grade is +0.31, although the uncrossed race of the same grade and generation gave offspring of mean grade about -1.20. The cross, therefore, had apparently increased the pigmentation of the extracted hooded recessives. This idea is supported by the result of a control mating of the particular female used in making the cross. When she was mated with a hooded male of the same grade as herself, she produced three young, all of grade -1.00. The extracted recessive grandchildren, as a group, show greatly increased pigmentation as compared with this, but vary greatly in the extent of the increase. Some show very little modification, others very much, the most extreme individual being of grade +3.50. It was undoubtedly out of just such modified recessive individuals as this that the material for our initial plus selections arose; to this point we shall return later.
The F₂ (or second generation) offspring, however, include about 1 hooded individual in 4. In a total of 962 F₂ young; 230 were hooded, or 24 per cent. This summary includes only those litters in which dominants as well as recessives were recorded. In many litters only the hooded young were recorded, as the special object of the investigation was to ascertain whether the extracted recessives were like the pure hooded race in grade or not. In the above summary also the hooded grandparent was in every case a female. The reciprocal cross is more difficult to obtain, but one wild female rat, caught in 1911, has bred quite regularly in captivity, though each time she has murdered her hooded mate prior to the birth of the young. Her F₂ grandchildren derived from matings with males of the minus series include 32 hooded and 96 non-hooded individuals, exactly 25 per cent hooded.
A second cross of selected animals of the minus series was made between a wild male and four females of grade -2 derived respectively from generations 5¼, 5½, 6½, and 7. As a group these mothers are more nearly comparable with generation 6, Table 21, than with any other uncrossed group. As the F₁ progeny of these four mothers by a wild male were mated inter se, it is possible to deal with their hooded grandchildren only as a group. The character of these is indicated in the second row of Table 42. They number 48 individuals and have a mean grade of +0.25, showing a modification in a plus direction similar to that observed in the previous case.
A third cross in which the minus series is concerned was made between females of grade -2 and -2.25, generation 10, and wild males. The F₂ offspring include 91 hooded individuals classified as to grade in the third row of Table 42. Their mean grade is +0.24, confirming fully the results obtained in the two previous experiments.
With these three cases we may compare three cases in which animals of the plus selection series were crossed with a wild male. (See the last three rows of Table 42.) Females of grade +3.00, generation 3, were crossed with a wild male. From this mating resulted 21 hooded grandchildren, ranging in grade from +1.75 to +3.50, mean +2.56. These grandchildren, it will be observed, in no case are of minus grade, as are about half the grandchildren when the grandparent is of minus grade. There is also no clear evidence of modification of the hooded character by the cross in this case. The grade of the extracted hooded individual is just about what uncrossed individuals of grade +3.00 produced in the corresponding generation of the plus series.
In the next case two females of the plus series, belonging to generations 5 and 6, respectively, were crossed with a wild male and their children were bred inter se. There resulted 38 hooded grandchildren, as shown in the next to the last row of Table 42. The range of the grades of these hooded grandchildren was similar to that of the grandchildren in the foregoing case, but their mean was somewhat higher, as[18] we should expect, since they are descended from more highly selected individuals; for the hooded grandparents in this case were of grade +3.25 (generation 5) and +3.50 (generation 6), whereas the grandparent in the foregoing case was of grade +3.00 (generation 3).
It is a noteworthy fact that in both these cases the wild cross does not seem to have increased the pigmentation in extracted hooded individuals, as it did when the minus series was crossed, but rather to have diminished it; yet the difference between observed and expected is not great. We might disregard it altogether, if a similar but more striking result were not observable in the third case as well as in another series of crosses presently to be described.
The third case (last row of Table 42) involves a cross between a female of grade +4.25, generation 10, plus series, and a wild male. The F₂ offspring include 16 hooded individuals of mean grade +3.15. Animals of this grade in the uncrossed race in this generation produced young of mean grade +3.84.
Before leaving this subject it is important to observe the considerable difference between the extracted hooded grandchildren of the minus-series rats, as a group, and those of the plus series. The latter is unmistakably a plus-series group; the former is on the border line between the two series. (Cf. Plates 2 and 3.)
As a control on the results given by the wild crosses, we may examine the results obtained by crossing the plus and minus selected races with a black Irish race. The Irish race used for this purpose consisted of animals black everywhere except on the belly. On the system of grading used in this paper they would range from +4½ to +5¾, +6 being an all-black rat.
Crosses of minus-series hooded rats with Irish produced Irish F₁ offspring with rather more white on the belly than the Irish parents possessed. In the F₂ generation hooded individuals reappeared in approximately the expected 25 per cent. In a total of 764 second generation young, 171, or 22.4 per cent, were hooded. The grade of pigmentation of these extracted recessives as compared with that of their hooded grandparents we may now consider, as was done in the case of the wild crosses. (See Table 43.)
Six individuals of the minus selection series, of generation 3½, and of mean grade -1.50, were crossed with Irish rats producing Irish offspring which were mated inter se. Among the grandchildren appeared the usual proportion of recessives (hooded), 90 in number. The distribution of these as regards grade of pigmentation is shown in the first row of Table 43. Their mean grade is -0.62, that of uncrossed hooded rats of the same grade as the hooded grandparents being -1.31 in generations[19] 3 and 4. In other words, the cross has considerably increased the pigmentation in the hooded grandchildren over what was to be expected had the cross not taken place. Nevertheless the increase in this case is less than in the similar cross with wild rats. (Compare Table 42.)
In the second row of Table 43 is shown the grade distribution of extracted hooded grandchildren of two mothers of grade -1.87 and generation 4. The mean of the 53 hooded grandchildren is in this case -0.73, that of uncrossed hooded parents of the same grade and generation being 1.18. This average is probably too low. An examination of the means of adjacent classes (Table 19) indicates that it should be about 1.35.
In the third row of Table 43 is shown the grade distribution of the extracted recessive grandchildren of a -2.00 male, minus-series rat, of generation 7½. The 66 grandchildren are of mean grade -0.94, expected -1.75.
Comparing the three experiments (first three rows of Table 43), we see that the more advanced grandparents, in grade and generations of selection, have the more advanced grandchildren; but in every case these are less advanced than grandparents of the same sort would have given had they not been crossed. Hence crossing with Irish has clearly had the effect of increasing the pigmentation in the minus series in the same way (but in lesser degree) as did crossing with wild animals.
The results of crossing hooded rats of the plus series with Irish ones are shown in the last two rows of Table 43. Several rats of mean grade +2.25 and of generation 2 were crossed with Irish, and their Irish young were then bred inter se, producing 239 hooded grandchildren. These ranged in grade from -1.00 to +3.25, their mean being +1.27. The grade of uncrossed rats of like grade and generation to the hooded grandparents is +1.80. Hence here, as in the cross with wild rats, the pigmentation has not been increased, but decreased by the cross, contrary to what we should expect. Further, the departure from expectation is greater in this cross than in the wild cross. These conclusions are supported by the results shown in the last row of Table 43. In the experiment here recorded a +3.00 rat of generation 3 was mated with an Irish rat. The hooded grandchildren derived from this cross were, as shown in the table, of mean grade +0.95, expected about +2.50. Since the number of animals recorded in this experiment is comparatively small, the quantitative result is less important than that of the foregoing experiment, but qualitatively the two are in entire agreement.
The various crosses of the selected minus and plus series with wild rats and with Irish rats respectively are consistent with each other. In every case the cross increases the pigmentation of the minus series and decreases that of the plus series; in other words, it undoes the work of selection to some extent. Does this mean that the condition created by selection was in reality an unstable one, so that an outcross tends to do[20] away with it? We do not think so, but to this question we shall return again.
The question might be asked whether the modifications produced in the selected races by a cross with wild or Irish stock are likely to be more or less permanent than those produced in unselected races by the same means. A single experiment was made which bears on this question in relation to the Irish cross. One of the -2.00 grandchildren recorded in the third row of Table 43 was mated with -2.00 individuals of the uncrossed stock of the minus series and produced nine young of mean grade -0.63, the expectation for the uncrossed race of the same grade and generation being about -1.90. In other words, this extracted -2.00 individual regressed (in breeding) as if it really had been affected by the cross, even though it did not show it, but the number of young is so small that no emphasis should be placed upon this result.
From the experiment recorded in the last row of Table 43 were obtained extracted individuals of mean grade +1.37, which as parents produced 16 young of mean grade +1.68, or, in other words, offspring about like themselves. Hence the changes effected by a cross are permanent, like those effected by selection.
It has been suggested that the original material out of which the plus series came consisted of modified individuals produced by a cross with the wild race. This was not known positively to be so, because part of the original stock (with which MacCurdy worked) consisted of hooded black and hooded gray rats captured in company with gray self and black Irish rats and albinos. Subsequent experiments showed that ordinary albino rats, if crossed with wild gray ones, will produce in F₂ all these classes of individuals. This indicated pretty clearly that the particular colony which had fallen into our hands had probably arisen by the crossing of an escaped albino rat with wild ones. But it still remained uncertain what sort of hooded pattern the escaped albino had transmitted and whether or not this had been influenced by the wild cross. We therefore determined to ascertain whether out of our minus series crossed with wild a plus series could be derived. To this end certain of the F₂ extracted hooded individuals (entered as grandchildren in Table 42, row 1, and descended from a single hooded individual of grade -1.87, generation 2½) were mated inter se, thus producing an F₃ generation, Table 44, second row. The selected individuals were the aberrant male of grade +3.50 and females of grade +1.50, so that the mean grade of the chosen parents (extracted from the crossed minus series) was +2.50. They had 34 young ranging in grade from 0 to +3.50, mean +2.06, a regression of 0.44 toward 0, repeating the phenomenon regularly found in both selection series.
In this same experiment some F₂ parents of mean grade -0.75 had 19 young (first row of Table 44), whose mean grade was -0.04, a[21] regression of 0.71 toward 0. We should expect the regression of the offspring of such parents to be less than that of the offspring of the -2.50 parents, and so it would be if it were not for one aberrant individual. Larger numbers of offspring would undoubtedly have given the expected result.
From among the F₃ offspring were chosen parents for the next generation (F₄). The chosen parents ranged in mean grade from +2.25 to +3.12 (Table 45), average +2.52. They produced 205 young ranging in grade from -0.25 to +3.50, mean +1.86, a regression of 0.66.
The parents for the next generation (Table 46) ranged in mean grade from +2.00 to +3.00, the mean being +2.27. They produced 119 offspring of mean grade +2.06, a regression of only 0.21.
| Selection. | Present series. | General (plus) series. | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean, parents. | Mean, offspring. | Regression. | No. of offspring. | Mean, parents. | Mean, offspring. | Regression. | No. of offspring. | |
| 1 | 2.50 | 2.06 | 0.44 | 34 | 2.51 | 2.05 | 0.46 | 150 |
| 2 | 2.52 | 1.86 | .66 | 205 | 2.52 | 1.92 | .60 | 471 |
| 3 | 2.27 | 2.06 | .21 | 119 | 2.73 | 2.51 | .22 | 341 |
| 4 | 2.69 | 2.41 | .28 | 194 | 3.09 | 2.73 | .36 | 444 |
| 5 | 2.77 | 2.32 | .45 | 97 | 3.33 | 2.90 | .43 | 610 |
| 6 | 3.08 | 2.67 | .41 | 45 | 3.52 | 3.11 | .41 | 861 |
The parents chosen from among these offspring ranged in mean grade from +2.37 to +3.25, average +2.69. They produced 194 offspring of grade +0.50 or higher (F₆, Table 47), the range for the first time lying wholly in the plus direction. The mean grade of the offspring was +2.41, a regression of 0.28.
The parents of the next generation (F₇, Table 48) range in mean grade from +2.62 to +3.37, their average being +2.80. Their 154 offspring range from +0.75 to +3.75, mean +2.46, a regression of 0.34.
The parents of the last generation in this experiment (F₈, Table 49) were of mean grade +3.08. They produced 45 offspring of mean grade +2.67, a regression of 0.41.
As a result of a single cross with a wild race followed by six successive selections, a narrow-striped or minus family has thus been converted into a wide-striped or plus family. Considering the smaller number of offspring from which selections could be made, progress was quite as rapid in this series as in the larger plus selection series. The regression is surprisingly similar, generation by generation, in the two series. (See Table B.) But it seems improbable that the closeness of the agreement has any significance. This series has the theoretical advantage of being derived from a single individual of the minus selection series.
When animals of the plus selection series are crossed with animals of the minus selection series, an F₁ generation of offspring is obtained which varies about a mean intermediate between those of the respective uncrossed races. Thus, from an examination of Table 50 it will be seen that when -2 animals of generation 6, minus series, were crossed with +3.50 or +3.75 animals of generation 5, plus series, an F₁ generation (Series 1) was obtained consisting of 93 animals of mean grade +0.06. This generation is rather more variable than either uncrossed race, its standard deviation being 0.71. The same is true of a second set (Series 2) of crosses made between a male of grade -3.25, generation 10, minus series, and females of grade +3.75, generation 10, plus series. The 14 F₁ offspring are of mean grade +1 and have a standard deviation of 0.60. (See Table 50, Series 2.)
In both the series of crosses summarized in Table 50, the F₂ generation is more variable than F₁. In Series 1, 305 F₂ animals are recorded, having a standard deviation of 1.01 as compared with 0.71, the standard deviation of the F₁ generation. In Series 2, the F₂ offspring number 73 and have a standard deviation of 0.87, that of the previous generation being 0.60.
The mean of the F₂ generation is very similar to that of the F₁ generation. In Series 1, the mean of F₁ is 0.06, and that of F₂ is 0.24; in Series 2, the mean of F₁ is 1.00, and that of F₂ is 0.72.
It may also be seen from an examination of Table 50 that among the F₁ offspring produced by crossing the plus and the minus series there are differences in transmission, as there are in the expression of the hooded pattern. In general those F₁ individuals which are of high grade produce offspring of higher grade than do their low-grade brothers and sisters. This is exactly what has been observed in both uncrossed races.
The experiments which have been described in the foregoing pages have shown that:
1. The hooded pattern of rats behaves as a simple Mendelian character in crosses with either the Irish pattern or the wholly pigmented condition of wild rats.
2. Though behaving as a unit, the hooded pattern fluctuates—that is, it is subject to plus and minus variations.
3. Selection, plus or minus, changes the position of the mean and mode about which variation occurs.
4. The results of such plus or minus selections are permanent, for return selection is not more effective than the original selection, and during return selection regression occurs away from the original mode, that is, toward the mode established by selection.
5. During the progress of the original selection (thirteen successive generations) variability as measured by the standard deviation was somewhat diminished.
6. Upon crossing the selected plus and minus races with each other, the variability was somewhat increased in F₁ and was further increased in F₂. The extreme conditions (plus or minus) of the grandparents rarely, if ever, recur in this generation. Only one individual among 378 F₂ young has been recorded in a grade as extreme as either grandparent.
7. Hooded animals extracted in F₂ as recessives from a cross with either Irish or wild rats are as a rule more variable than the selected race used in making the cross. In crosses with an Irish race the minus series was affected in like measure. In crosses with wild rats the variability of the plus series was not appreciably affected (in two experiments it was slightly reduced, and in one experiment it was slightly increased). But the variability of the minus race was more than doubled by crosses with wild rats.
8. The mean of the minus race was lowered by a cross with either the Irish race or with wild rats, but more extensively by the latter. The mean of the plus race was lowered a very little by a cross with wild rats, but considerably by a cross with the Irish race.
It would be possible to suppose, as the senior author has elsewhere suggested (Castle, 1912), that the Mendelian unit character involved in these experiments is subject to quantitative variation and that such quantitative variations have a tendency to persist from generation to generation. This would account for the effectiveness and permanency of selection when brought to bear upon the variations. It might also form a basis for explaining the increased variability which follows crossing, this being regarded as due to contamination in the heterozygote, but there are certain other observed effects of crossing which it seems impossible to account for on this basis. In particular it is observed that while crossing the minus series makes it less minus as the hypothesis of contamination would demand, crossing the plus series makes it less plus, the opposite of what a contamination theory would demand. For we can readily understand, on the basis of contamination, how a +6 gamete being combined with a -2 gamete might change the latter in a plus direction; but if the same +6 gamete is associated with a +4 gamete we should expect it, if it has any influence at all, to make this also more plus, but the observed effect is the opposite; the extracted gametes are less plus in character.
This difficulty is met by an alternative explanation, the main feature of which was first suggested by our colleague, Dr. E. M. East, viz, that although we seem to be dealing with a single unit character as evidenced by the monohybrid ratios obtained, nevertheless the modifications[24] which form a basis for selection are due (in part at least) to agencies transmitted independently of the hooded pattern (not forming a part of the same unit character), and which may be present in Irish as well as in wild rats. By crosses with such rats the supposed modifiers may become associated with the hooded pattern in extracted recessive individuals and so increase its extent. Such increase does actually occur in experiment.
The hypothesis of modifiers independent in transmission of the hooded unit will account for the fact that F₂ is more variable than F₁ when crosses are made, on the familiar principle of recombination of independent factors. It will account for the observed effectiveness of selection on the ground that what selection accomplishes in the plus series is the isolation of homozygous conditions of modifiers at first present only in heterozygous form, and that what it accomplishes in the minus series is the isolation of conditions homozygous for lack of modifiers (or for inhibitors) of pigmentation. This same hypothesis will account also for the observed reduction of variability during the progress of selection, for as soon as any particular modifier attains a homozygous condition in the race it will cease to occasion variability, and as more and more factors become homozygous the variability should accordingly diminish and finally disappear altogether, so far as it is due to internal and heritable causes.
At this point the hypothesis of modifiers encounters serious difficulty, if one holds the prevalent or “genotype” conception as to the nature of Mendelian factors, viz, that they are fixed and unchangeable and not subject to quantitative variation, but only to combination in different ways with other factors. This conception has been presented very clearly by Dr. East (1912). Some objections to this view had previously been stated by Castle (1912) and need not here be repeated.
If we assume that there exists at the outset a definite number of modifiers and that these possess a definite and unchanging power to modify, then it is evident that selection can do nothing but secure homozygous conditions as regards the presence or absence of these modifiers. When such homozygous conditions are secured, selection will cease to modify the race. The experiment has progressed far enough to show that extensive modification through selection is possible without any marked falling off in variability. No indication is observable that selection will become ineffective before an all-black rat is obtained in the plus series and an all-white rat in the minus series. A nearly all-black race of rats has already been secured. We propose to continue the experiments until demonstrative evidence is obtained.
If the fixed-factor idea as regards modifiers of the hooded pattern is rejected, there remain still two possible alternative views regarding them. Either we may consider that the modifiers vary in strength, that is, in power to modify, or we may consider that new modifiers arise[25] from time to time, which selection may either add in homozygous form to the germinal complex or reject altogether from it. If we assume that the modifiers vary in strength, we shall have to grant also the possibility that the character modified, the hooded pattern, may itself vary in strength independently of its modifiers. For evidence see the description of the “mutant” series, page 30. This assumption, I understand, would be unacceptable to those who hold a genotype conception of heredity, though we ourselves can offer no valid objection to it.
If, on the other hand, we admit that new modifiers or inhibitors are from time to time coming into existence spontaneously, and that selection can use these to modify the pattern either in a plus or in a minus direction, then we must admit that selection is an agency of real creative power, able to modify unit characters indefinitely so long as physiological limitations are not reached.
Now it seems to us probable that what we call the unit-character for hooded pattern is itself variable; also that “modifiers” exist—that is, the extent of the hooded pattern is not controlled exclusively by a single localized portion of the germ-cell; otherwise we should be at a loss for an explanation of the peculiar results from crossing plus series hooded rats with those which are still more extensively pigmented; for by such crosses the pigmentation is rendered not more extensive but less so. This result we can explain on the supposition that the selected plus series has accumulated more modifiers of the hooded pattern than the wild race contains, so that a cross tends to reduce the number of modifiers in the extracted hooded individuals. No other explanation at present offers itself for this wholly unexpected but indubitable result. If a different one can be found we are quite ready to discard the hypothetical modifiers as a needless complication, contenting ourselves with the supposition that the unit character for hooded pattern is itself variable, and that for this reason racial change in either plus or minus directions may be secured at will through repeated selection.
We have been led to adopt tentatively an hypothesis that modifying factors exist independent of the single factor for hooded pattern (though both the factor for hooded pattern and its modifiers may, so far as we can see, be quantitatively variable) by another series of observations, which will now be described.
In the tenth generation of the plus selection series there appeared two individuals, a male and a female, of considerably higher grade than any previously recorded in this series. They are not included in Table 10 because we have been and still are in doubt as to their exact nature and think it best to give a separate account of them. If entered in Table 10 one would appear as a 5½ individual born of 3⅞ parents (mean grade), the other as a 5¾ individual born of 3¾ parents (mean grade). The[26] nearest individuals in grade to these two produced by the same group of parents are of grade 4½, but some 4⅛ parents of the same generation produced two offspring of grade 5. (See Table 10.) Because of the marked advance in grade of these individuals beyond the ordinary range of variation in the series we called them “mutants,” without wishing then or now to commit ourselves to any particular theory as to their nature or origin. We have used the term and now use it as one of convenience merely. The two “mutant” individuals had the same father and their mothers were sisters. Their pedigree for two generations is as follows:
| Mutants. | Parents. | Grandparents. | |||
| ♂ 4763, +5½ | { | ♀ 2956, +3¾ | } | { | ♀ 1939, +3¾ |
| { | ♀ 2957, +3½ | } | { | ♂ 1817, +3¾ | |
| { | |||||
| ♀ 5153, +5¾ | { | { | ♀ 1162, +3½ | ||
| { | ♂ 2963, +4 | { | ♂ 1810, +3¾ | ||
The mutant male was mated with the mutant female and also with other females of the plus series, with the results shown in Table 51. In every case the young fall into two distinct groups, one of which varies about the general mean of the plus series (approximately 3¾), while the other varies about the father’s grade as a mean (approximately 5½).
The mutant female had 16 young, 6 in the lower group, mean 3.87, and 10 in the upper group, mean 5.60. (See Table 51, lowest row.) The other females had in all 114 young almost equally divided between the two groups, 58 in the lower group, mean 3.73, and 56 in the upper group, mean 5.45. This result indicates clearly (what the sequel also confirms) that the male mutant transmitted in half his gametes the high grade of pigmentation which he himself manifested, while in the other half of his gametes he transmitted the ordinary condition of the plus race at that time. In other words his “mutant” character behaved as a dominant unit in relation to the ordinary condition of the plus race.
It is evident that the female mutant was of similar constitution. This being the case, we should expect three-fourths of the offspring of the two mutants to be in the upper group. In reality 10 of their 16 young were of this sort.
The male mutant was mated also with females of the minus series with the results indicated in Table 52. Again, the offspring fall into two distinct groups, a lower and an upper. The lower group should be comparable with the result obtained in F₁ when the plus and minus races are crossed with each other. (Compare Table 50.) Such it proves to be. It includes 35 individuals of mean grade -0.49 and standard deviation 0.77. Series 2 of Table 50 is nearly contemporaneous with this experiment. The F₁ offspring in that series were of mean grade -1 and standard deviation 0.60.
The upper group of offspring (Table 52) result, we may suppose, from a mutant gamete (grade about 5½) united with a narrow series gamete (grade about -2). This group includes 31 individuals varying closely about grade 4½, and with a standard deviation of only 0.31. The lower average grade of this group (4.43) compared with the similar group of Table 51, which had a mean of 5.47, shows the influence of the minus-series gamete upon the heterozygote in lowering its grade by about 1. Whether the plus-series gametes have any effect upon the grade of the heterozygotes recorded in the upper group of Table 51 is not certain, because a homozygous group of mutants has not yet been established. It may be observed, however, that one individual in the upper group of Table 51 was of grade 6 (colored all over), and it is possible that homozygous “mutants,” when obtained, will approximate that grade, as most wild rats do. Further, a comparison of Tables 51 and 53 shows that mutant heterozygotes formed by crosses with the plus series are of slightly lower mean grade than the offspring of the two mutants, among which should occur both homozygous and heterozygous mutants. It seems probable, therefore, that homozygous mutants will be found to be of somewhat higher grade than heterozygous ones.
The question early suggested itself to our minds, will these “mutants” prove to be mutants in the sense of De Vries? Will they prove to be more stable than the modifications ordinarily secured by selection in our experiments? To test this matter, we have raised two additional generations of offspring from the two mutants and have bred a second generation of offspring from each of the four groups of F₁ offspring recorded in Tables 51 and 52, derived from matings with the plus and minus races respectively.
The F₂ descendants of the two original mutants proved very similar to the F₁ descendants. (See Table 53.) They fall as before into two groups, an upper and a lower. The former includes 30 individuals of mean grade 5.52, the latter 2 of mean grade 3.37. As the parents of this generation were taken wholly from the upper group of offspring of generation F₁, and as theoretically that group should contain 2 heterozygous individuals to one which is homozygous for the “mutant” character, it is to be expected that in F₂ more than three-fourths of the offspring will fall in the upper group. For any pair, one member of which is homozygous for the mutant character, should produce only offspring falling in the upper group; and offspring falling in the lower group should be produced only by pairs both members of which are heterozygous.
The upper group in F₂ should contain a larger proportion of homozygous mutants than in F₁, and since the parents of F₃ were chosen from this upper group of F₂ offspring, it is not surprising that the 11 F₃ offspring recorded up to this time all fall in the upper group. The mean of this upper group is remarkably constant through the three generations,[28] and the variability of the group as measured by its standard deviation is also low, namely, 0.19. This indicates that the mutant character is a strongly dominant unit in relation to the ordinary condition of the plus series.
Table 54 shows the character of the F₂ offspring of the original male mutant mated with females of the plus series. The lower group parents, those into which the mutant character did not presumably enter at all, produced 59 offspring recorded in the first part of Table 54. Their mean grade is 3.78 and their standard deviation 0.33. These are very close to the constants of the general plus series, which for generation 10 were 3.73 and 0.36, respectively.
The second division of Table 54 shows the character of the young produced by the F₁ parents of the upper group (Table 51). Such parents are supposed to have received a “mutant” gamete from their father, grade about 5.50, and a plus-series gamete from their mother, grade about 3.75. If they produce gametes of these same two sorts, their offspring should also fall into two corresponding groups; in fact they do. There are 11 offspring of mean grade 3.86 and 79 offspring of mean grade 5.50. As in the previous generation, the two groups do not approach each other in grade. The mean and standard deviation of the lower group of offspring are similar to those of the plus race. The mean of the upper group is about the same as that of their parents (upper group of offspring, Table 51), namely, 5.50, as compared with 5.45; their standard deviation is somewhat lower, namely, 0.15, as compared with 0.23. This result indicates that the “mutant” character and the hooded character of the plus series segregate from each other in a simple way without modifying each other appreciably. It seems possible that they contain the same modifiers (if modifiers are present) and differ merely by the main unit which we called the hooded character in the early part of this paper. Each contains a different condition of that main unit. Consequently there is no increase of variability in F₂ when these two conditions are intercrossed. This we should expect to happen, if they differed by more than a single factor.
A very different result is obtained from the cross between the mutant and narrow races. Although F₁ from that cross was quite variable (see Table 52), F₂ is still more variable (see Table 55). The lower group F₁ individuals, which resembled F₁ between the plus and minus races, produced 61 young (first division of Table 55), which resemble F₂ between the plus and minus races. They range in grade from -2 to +3¼, mean +0.58, standard deviation 1.17. In the two series of crosses between the plus and minus races (Table 50) the means were +0.24 and +0.72, respectively, and the standard deviations 1.01 and 0.87. This indicates, as did the cross with the plus series, that the “lower group” gametes produced by the original mutant male did not differ materially from gametes produced by the ordinary plus race from which the mutant sprang.
The second division of Table 55 shows the character of the F₂ young produced by the upper group of F₁ offspring recorded in Table 52. It consists of two groups, a lower and an upper. The lower represents the extracted minus race, the upper represents the extracted dominants or mutants, whether homozygous or heterozygous. The former group has an average of +0.75 and a standard deviation of 1.03, which values are close to the corresponding constants of Series 2, Table 50, the latest of the plus-minus crosses, in which the mean was +0.72 and the standard deviation 0.87.
The upper group offspring of Table 55, second division, the homozygous and heterozygous mutants, number 68; they have a mean grade of 4.77 as compared with 4.43 in F₁, which consisted exclusively of heterozygotes. This shows the extracted homozygotes to be of higher grade than the heterozygotes. The highest grade mutant among the 31 F₁ young, all of which were heterozygotes, was of grade 5, but among the 68 F₂ young are 16 of higher grade than 5. We expect one-third of these 68 individuals to be homozygotes. Now all of the F₂ mutants from the cross of mutant with plus race (Table 54) were of grade 5 or higher, only 2 in 79 being as low as 5, and 13 of the 79 being of grade 5¾, a grade not attained at all in F₂ from the mutant-minus cross (Table 55). This result shows us that the cross with the minus race does affect permanently the mutant character, lowering its grade even in homozygous mutants extracted from the cross. It also increases the variability of the mutants, for the standard deviation of the mutant group in Table 55 is 0.44, whereas in Table 54 (mutant-plus F₂), in a like number of individuals, it was 0.15, or only about one-third as great.
That the variability of the mutants is unaffected by a cross with the plus race, but that it is increased by a cross with the minus race, and that, further, the mean of the mutants is affected little or none by a cross with the plus race, but that it is lowered by a cross with the minus race—these several facts are all conformable with the hypothesis that the change in variability due either to crossing or to selection results from modifying factors which, as they are independent of the main factor concerned, are probably transmitted in a different part or component of the germ-cell than that factor. For if the mutant and the plus race are alike as regards the modifiers, but differ only in the main factor, then no change in variability should result from intercrossing them, but only alternative conditions as regards the main factor. This is the observed result. But if the mutant and the minus race differ not only in the main factor, but also in modifiers which are independent of it, then, when they are crossed, we may expect that through independent segregation of main factor and modifiers the extracted minus race will be raised in grade, while the extracted mutants are lowered, and both will become more variable. This also is the observed result.
One objection may be offered to this interpretation, namely, that the increased variability is not delayed until F₂, but is already in evidence[30] to some extent in F₁. The same thing was observable in the crosses of the plus and minus series (Table 50). From that table, Series 1, it will be observed that when the plus and minus races had standard deviations of 0.49 and 0.50, respectively, their F₁ offspring had a standard deviation of 0.71, an increase by nearly one-half; F₂ showed a further increase to 1.01. In series 2, Table 50, the uncrossed races (generation 10) had standard deviations of 0.36 and 0.24; their F₁ offspring had a standard deviation of practically twice this, namely 0.60; F₂ showed a further increase to 0.87.
At the time of the mutant-minus race crosses, the minus race (generation 10) had a standard deviation of 0.24, the plus race of 0.36. F₁ (lower group) had a standard deviation of 0.77, and F₂ of 1.17. F₁ mutants (upper group) had a standard deviation of 0.31 which rose in F₂ to 0.44. These various facts will perhaps be better grasped if presented in tabular form:
| Standard deviation of races crossed, same generation. |
S. D. F₁ | S. D. F₂ | ||
|---|---|---|---|---|
| Plus-minus cross, series 1 (Table 50) | 0.49 | 0.50 | 0.71 | 1.01 |
| Plus-minus cross, series 2 (Table 50) | .36 | .24 | .60 | .87 |
| Mutant-minus cross, lower group | .25 | .24 | .77 | 1.17 |
| Mutant-minus cross, upper group | .19 | .24 | .31 | .44 |
| Mutant-plus cross, lower group | .25 | .36 | .24 | .35 |
| Mutant-plus cross, upper group | .19 | .36 | .23 | .15 |
The mutant-plus cross, it will be observed, shows no increase of variability either in F₁ or in F₂, but crosses involving the minus race show increase of variability both in F₁ and in F₂. Interpreted on a Mendelian basis, this means that the mutant and plus races on the one hand and the minus race on the other hand differ by more than a single factor. If they differed by only a single factor, then crosses between them should bring no increase of variability, either in F₁ or in F₂. This appears to be true as regards the mutant and plus races when crossed with each other. But if the races crossed differ by more than one factor, and if, further, neither parent is homozygous as regards the factors in which they differ, then we may expect an increase in variability both in F₁ and in F₂. This is exactly what we observe when the minus race is crossed with either the plus race or its derivative, the mutant race.
If we suppose that the plus race and the minus race differ from each other by certain “modifiers,” we can not suppose that the plus and the mutant races differ by these same modifiers. They differ in some other single respect; perhaps that in which they differ is the main hooded factor. Are we, then, to suppose that the plus and the minus races do not differ as regards this same main factor? This can not be stated, but[31] we see no reason for considering them identical as regards that factor. It appears that the mutant race arose from the plus race by a single large plus variation, which seems to have its determiner in some single component of the germ-cell. But the fact that this change came as a large quantitative variation does not show that small variations are impossible in that same cell component. It seems to us quite improbable that the plus mutation could have arisen in the minus selection series. We believe that the repeated selection which was practised had something to do with inducing this change in the plus direction. If one can increase at will the “modifiers” which make the pigmentation more extensive, it does not seem strange that after a time a readjustment should occur within the cell which should incorporate modifiers in that part of the cell which is responsible for the unit-character behavior of the hooded pattern. This would amount to a quantitative change in the unit-character for hooded pigmentation.
1905. Heredity of coat characters in guinea-pigs and rabbits. Carn. Inst. Wash. Pub. 23.
1906. The origin of a polydactylous race of guinea-pigs. Carn. Inst. Wash. Pub. 49.
1912. The inconstancy of unit-characters. American Naturalist, vol. 46, pp. 352-362.
1906. Heredity of hair-length in guinea-pigs and its bearing on the theory of pure gametes. Carn. Inst. Wash. Pub. 49.
1901-1903. Die Mutationstheorie. Veit & Co., Leipzig.
1912. The mendelian notation as a description of physiological facts. American Naturalist, vol. 46, pp. 633-655.
1909. Heredity and variation in the simplest organisms. American Naturalist, vol. 43, pp. 321-337.
1910. Experimental evidence on the effectiveness of selection. American Naturalist, vol. 44, pp. 136-145.
1909. Elemente der exakten Erblichkeitslehre. G. Fischer, Jena.
1907. Selection and cross-breeding in relation to the inheritance of coat-pigments and coat-patterns in rats and guinea-pigs. Carn. Inst. Wash. Pub. 70.
1913. Genetics and breeding. Science, n. s., vol. 37, pp. 539-546.
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | ||||
| 1⅞ | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 7 | 1.82 | .05 | ||
| 2 | 4 | 1 | 3 | 1 | 4 | 1 | 4 | 18 | 1.76 | .24 | ||
| 2⅛ | ||||||||||||
| 2¼ | 3 | 1 | 3 | 1 | 6 | 2 | 2 | 2 | 20 | 1.87 | .38 | |
| 2⅜ | ||||||||||||
| 2½ | 5 | 3 | 1 | 13 | 1 | 8 | 5 | 1 | 37 | 2.06 | .44 | |
| 2⅝ | 1 | 1 | 2 | 1 | 5 | 2.15 | .47 | |||||
| 2¾ | 7 | 3 | 2 | 17 | 1 | 12 | 9 | 51 | 2.12 | .63 | ||
| 2⅞ | ||||||||||||
| 3 | 2 | 3 | 1 | 3 | 3 | 12 | 2.35 | .65 | ||||
| Totals or means, 2.51 | 20 | 3 | 13 | 8 | 44 | 7 | 31 | 10 | 4 | 150 | 2.05 | .46 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -1 | -¾ | -½ | -¼ | 0 | +¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | ||||
| 2 | 1 | 1 | 4 | 2 | 5 | 7 | 3 | 7 | 8 | 2 | 3 | 2 | 1 | 46 | 1.70 | .30 | |||||||
| 2⅛ | 1 | 1 | 2 | 1 | 3 | 5 | 12 | 8 | 5 | 5 | 8 | 2 | 1 | 2 | 56 | 1.28 | .82 | ||||||
| 2¼ | 1 | 1 | 3 | 1 | 1 | 1 | 8 | 1.87 | .38 | ||||||||||||||
| 2⅜ | 1 | 1 | 4 | 9 | 7 | 8 | 8 | 6 | 1 | 45 | 1.92 | .45 | |||||||||||
| 2½ | 4 | 22 | 16 | 19 | 11 | 21 | 13 | 7 | 10 | 9 | 1 | 133 | 1.80 | .70 | |||||||||
| 2⅝ | 1 | 2 | 3 | 1 | 6 | 5 | 7 | 9 | 6 | 3 | 1 | 44 | 2.11 | .51 | |||||||||
| 2¾ | 1 | 3 | 4 | 2 | 6 | 5 | 9 | 6 | 9 | 6 | 1 | 52 | 1.92 | .83 | |||||||||
| 2⅞ | 1 | 1 | 3 | 5 | 3 | 9 | 1 | 23 | 2.41 | .46 | |||||||||||||
| 3 | 1 | 2 | 3 | 2 | 14 | 7 | 4 | 5 | 11 | 6 | 3 | 1 | 59 | 2.47 | .53 | ||||||||
| 3½ | 2 | 1 | 2 | 5 | 2.50 | .62 | |||||||||||||||||
| Totals or means, 2.52 | 1 | - | 2 | 1 | 4 | 1 | 5 | 16 | 45 | 37 | 45 | 45 | 77 | 57 | 44 | 48 | 30 | 9 | 3 | 1 | 471 | 1.92 | .60 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | ||||
| 2⅛ | 1 | 1 | 5 | 6 | 4 | 1 | 3 | 21 | 2.06 | .06 | |||||||
| 2¼ | |||||||||||||||||
| 2⅜ | 5 | 5 | 1 | 4 | 2 | 17 | 2.15 | .22 | |||||||||
| 2½ | 1 | 2 | 1 | 2 | 5 | 5 | 10 | 10 | 9 | 7 | 2 | 54 | 2.32 | .18 | |||
| 2⅝ | 2 | 1 | 3 | 9 | 4 | 5 | 15 | 18 | 3 | 1 | 2 | 1 | 64 | 2.64 | -.02 | ||
| 2¾ | 2 | 1 | 7 | 10 | 7 | 7 | 7 | 1 | 1 | 43 | 2.46 | .29 | |||||
| 2⅞ | 1 | 7 | 3 | 7 | 9 | 10 | 9 | 1 | 47 | 2.70 | .17 | ||||||
| 3 | 2 | 1 | 12 | 14 | 11 | 17 | 11 | 3 | 71 | 2.49 | .51 | ||||||
| 3⅛ | 2 | 5 | 1 | 2 | 10 | 3.07 | .05 | ||||||||||
| 3¼ | 1 | 2 | 1 | 2 | 6 | 3.17 | .08 | ||||||||||
| 3⅜ | 2 | 1 | 3 | 2 | 8 | 2.91 | .46 | ||||||||||
| Totals or means, 2.73 | 1 | 2 | 5 | 8 | 20 | 51 | 46 | 47 | 66 | 63 | 21 | 6 | 4 | 1 | 341 | 2.51 | .22 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | ||||
| 2½ | 2 | 2 | 4 | 3 | 11 | 2.55 | .05 | |||||||||
| 2⅝ | ||||||||||||||||
| 2¾ | 3 | 3 | 2 | 3 | 4 | 3 | 18 | 2.65 | .10 | |||||||
| 2⅞ | 1 | 3 | 3 | 7 | 15 | 4 | 3 | 2 | 37 | 2.97 | .10 | |||||
| 3 | 1 | 1 | 7 | 12 | 25 | 19 | 35 | 29 | 9 | 4 | 1 | 143 | 2.60 | .40 | ||
| 3⅛ | 1 | 2 | 11 | 14 | 21 | 28 | 30 | 11 | 2 | 2 | 122 | 2.69 | .43 | |||
| 3¼ | 1 | 1 | 6 | 8 | 13 | 17 | 9 | 8 | 1 | 64 | 2.89 | .36 | ||||
| 3⅜ | 1 | 6 | 2 | 3 | 9 | 2 | 23 | 2.70 | .67 | |||||||
| 3½ | 3 | 1 | 2 | 3 | 6 | 2 | 3 | 20 | 3.02 | .48 | ||||||
| 3⅝ | ||||||||||||||||
| 3¾ | 3 | 1 | 1 | 1 | 6 | 2.75 | 1.00 | |||||||||
| Totals or means, 3.09 | 1 | 1 | 1 | 2 | 12 | 30 | 60 | 58 | 96 | 110 | 45 | 19 | 9 | 444 | 2.73 | .36 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | ||||
| 2¾ | 1 | 2 | 4 | 4 | 1 | 12 | 3.00 | -.25 | ||||||||||
| 2⅞ | 4 | 4 | 4 | 2 | 3 | 2 | 2 | 21 | 2.87 | 0 | ||||||||
| 3 | 1 | 5 | 3 | 2 | 3 | 1 | 15 | 2.81 | .19 | |||||||||
| 3⅛ | 1 | 1 | 1 | 1 | 8 | 9 | 14 | 20 | 29 | 18 | 8 | 3 | 1 | 114 | 2.81 | .31 | ||
| 3¼ | 1 | 1 | 1 | 4 | 12 | 6 | 14 | 24 | 39 | 27 | 7 | 1 | 1 | 138 | 2.81 | .44 | ||
| 3⅜ | 1 | 1 | 5 | 14 | 11 | 25 | 36 | 28 | 15 | 7 | 2 | 145 | 2.94 | .43 | ||||
| 3½ | 6 | 5 | 10 | 14 | 16 | 8 | 8 | 3 | 69 | 2.86 | .64 | |||||||
| 3⅝ | 3 | 3 | 5 | 7 | 16 | 12 | 11 | 4 | 3 | 64 | 3.08 | .54 | ||||||
| 3¾ | 2 | 2 | 4 | 3 | 2 | 1 | 14 | 3.07 | .68 | |||||||||
| 3⅞ | 1 | 1 | 1 | 1 | 2 | 2 | 8 | 3.35 | .52 | |||||||||
| 4 | 1 | 1 | 1 | 1 | 2 | 1 | 7 | 3.36 | .64 | |||||||||
| 4⅛ | 1 | 1 | 1 | 3 | 3.00 | 1.12 | ||||||||||||
| Totals or means, 3.33 | 1 | 2 | 2 | 2 | 6 | 35 | 44 | 66 | 101 | 151 | 107 | 57 | 24 | 11 | 1 | 610 | 2.90 | .43 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | ||||
| 3 | 1 | 1 | 2 | |||||||||||||
| 3⅛ | 3 | 4 | 1 | 1 | 4 | 7 | 5 | 3 | 28 | 2.84 | .28 | |||||
| 3¼ | 1 | 3 | 3 | 8 | 26 | 31 | 31 | 24 | 15 | 1 | 143 | 3.10 | .15 | |||
| 3⅜ | 1 | 5 | 6 | 13 | 20 | 28 | 30 | 14 | 5 | 123 | 2.96 | .41 | ||||
| 3½ | 1 | 9 | 4 | 15 | 29 | 49 | 41 | 43 | 17 | 4 | 212 | 3.10 | .40 | |||
| 3⅝ | 4 | 8 | 11 | 24 | 34 | 36 | 37 | 21 | 5 | 1 | 181 | 3.16 | .46 | |||
| 3¾ | 1 | 1 | 2 | 1 | 3 | 4 | 18 | 13 | 20 | 13 | 2 | 78 | 3.22 | .53 | ||
| 3⅞ | 4 | 1 | 3 | 5 | 14 | 15 | 20 | 14 | 4 | 80 | 3.26 | .61 | ||||
| 4 | 1 | 2 | 1 | 5 | 4 | 1 | 14 | 3.41 | .59 | |||||||
| Totals or means, 3.52 | 3 | 5 | 32 | 24 | 54 | 113 | 183 | 172 | 166 | 90 | 17 | 1 | 861 | 3.11 | .41 | |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | 4¾ | ||||
| 3⅛ | 2 | 2 | 4 | 5 | 7 | 4 | 3 | 1 | 28 | 2.87 | .25 | ||||||
| 3¼ | 1 | 6 | 11 | 7 | 11 | 30 | 19 | 22 | 23 | 1 | 131 | 3.09 | .16 | ||||
| 3⅜ | 1 | 2 | 10 | 18 | 17 | 21 | 27 | 18 | 21 | 20 | 4 | 1 | 160 | 2.97 | .40 | ||
| 3½ | 6 | 9 | 9 | 21 | 31 | 28 | 35 | 34 | 3 | 1 | 177 | 3.18 | .32 | ||||
| 3⅝ | 7 | 11 | 13 | 26 | 47 | 53 | 75 | 49 | 7 | 1 | 289 | 3.23 | .39 | ||||
| 3¾ | 2 | 3 | 3 | 16 | 28 | 25 | 58 | 42 | 4 | 3 | 184 | 3.35 | .40 | ||||
| 3⅞ | 2 | 1 | 2 | 4 | 10 | 12 | 23 | 21 | 6 | 6 | 2 | 1 | 90 | 3.49 | .38 | ||
| 4 | 1 | 1 | 3 | 2 | 4 | 3 | 1 | 15 | 3.53 | .47 | |||||||
| 4⅛ | 1 | 1 | 1 | 3 | 3.75 | .50 | |||||||||||
| Totals or means, 3.56 | 2 | 2 | 35 | 55 | 56 | 105 | 183 | 159 | 240 | 195 | 29 | 12 | 2 | 2 | 1077 | 3.20 | .36 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | ||||
| 3¼ | 2 | 2 | 4 | 3.25 | 0 | ||||||||||
| 3⅜ | |||||||||||||||
| 3½ | 1 | 4 | 6 | 9 | 18 | 17 | 2 | 2 | 59 | 3.46 | .04 | ||||
| 3⅝ | 1 | 2 | 9 | 13 | 30 | 69 | 44 | 144 | 149 | 22 | 1 | 484 | 3.50 | .12 | |
| 3¾ | 2 | 1 | 9 | 12 | 55 | 39 | 152 | 173 | 23 | 2 | 469 | 3.49 | .26 | ||
| 3⅞ | 1 | 2 | 4 | 23 | 18 | 98 | 69 | 19 | 2 | 1 | 238 | 3.53 | .34 | ||
| 4 | 1 | 2 | 3 | 10 | 6 | 34 | 6 | 1 | 1 | 64 | 3.31 | .69 | |||
| 4⅛ | 2 | 5 | 2 | 14 | 13 | 14 | 10 | 60 | 3.72 | .40 | |||||
| 4¼ | 1 | 7 | 11 | 2 | 1 | 22 | 3.69 | .56 | |||||||
| 4⅜ | 2 | 5 | 1 | 8 | 3.96 | .41 | |||||||||
| Totals or means, 3.75 | 1 | 5 | 13 | 25 | 55 | 170 | 119 | 469 | 440 | 88 | 20 | 1 | 1408 | 3.48 | .27 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | ||||
| 3½ | 1 | 1 | 1 | 1 | 4 | 3.25 | .25 | ||||||||
| 3⅝ | 1 | 1 | 6 | 12 | 28 | 26 | 96 | 63 | 10 | 1 | 244 | 3.43 | .19 | ||
| 3¾ | 1 | 2 | 4 | 7 | 19 | 45 | 50 | 224 | 212 | 25 | 2 | 591 | 3.50 | .25 | |
| 3⅞ | 2 | 5 | 25 | 32 | 105 | 177 | 64 | 8 | 6 | 424 | 3.65 | .22 | |||
| 4 | 1 | 2 | 5 | 14 | 15 | 7 | 1 | 45 | 3.57 | .43 | |||||
| 4⅛ | 2 | 4 | 1 | 7 | 3.57 | .55 | |||||||||
| 4¼ | 1 | 1 | 2 | 4 | 3.75 | .50 | |||||||||
| 4⅜ | 3 | 3 | 3.50 | .87 | |||||||||||
| Totals or means, 3.78 | 1 | 3 | 5 | 17 | 38 | 105 | 110 | 443 | 473 | 109 | 11 | 7 | 1322 | 3.54 | .24 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | 5 | ||||
| 3¾ | 1 | 1 | 9 | 13 | 63 | 117 | 44 | 8 | 1 | 257 | 3.69 | .06 | ||
| 3⅞ | 1 | 3 | 3 | 16 | 95 | 142 | 62 | 20 | 5 | 347 | 3.72 | .15 | ||
| 4 | 4 | 26 | 61 | 20 | 7 | 2 | 120 | 3.76 | .24 | |||||
| 4⅛ | 11 | 17 | 7 | 6 | 1 | 2 | 44 | 3.87 | .25 | |||||
| 4¼ | 1 | 4 | 2 | 1 | 8 | 3.84 | .41 | |||||||
| Totals or means, 3.88 | 1 | 1 | 4 | 12 | 33 | 196 | 341 | 135 | 42 | 9 | 2 | 776 | 3.73 | .15 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | 4¾ | 5 | ||||
| 3¾ | 2 | 7 | 2 | 11 | 3.75 | 0 | |||||||
| 3⅞ | 2 | 7 | 17 | 87 | 162 | 41 | 11 | 3 | 3 | 333 | 3.70 | .17 | |
| 4 | 3 | 2 | 25 | 87 | 65 | 24 | 6 | 1 | 1 | 214 | 3.87 | .13 | |
| 4⅛ | 3 | 3 | 16 | 49 | 27 | 8 | 2 | 2 | 110 | 3.81 | .31 | ||
| 4¼ | 2 | 13 | 5 | 3 | 1 | 1 | 25 | 3.91 | .34 | ||||
| 4⅜ | 1 | 3 | 4 | 3.94 | .43 | ||||||||
| Totals or means, 3.97 | 2 | 13 | 22 | 132 | 319 | 143 | 46 | 12 | 7 | 1 | 697 | 3.78 | .19 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2½ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | 4¾ | 5 | 5¼ | ||||
| 3¾ | 4 | 20 | 7 | 4 | 35 | 3.83 | -.08 | |||||||
| 3⅞ | 2 | 19 | 17 | 6 | 2 | 2 | 48 | 3.96 | .09 | |||||
| 4 | 1 | 1 | 10 | 53 | 59 | 12 | 5 | 1 | 1 | 2 | 145 | 3.93 | .07 | |
| 4⅛ | 1 | 2 | 17 | 86 | 83 | 28 | 6 | 3 | 1 | 227 | 3.91 | .21 | ||
| 4¼ | 1 | 7 | 19 | 29 | 11 | 3 | 1 | 71 | 3.94 | .31 | ||||
| 4⅜ | 3 | 6 | 14 | 10 | 7 | 3 | 1 | 1 | 45 | 4.13 | .24 | |||
| 4½ | 1 | 4 | 1 | 1 | 7 | 4.14 | .36 | |||||||
| 4⅝ | 1 | 3 | 4 | 3 | 11 | 3.90 | .72 | |||||||
| 5 | 1 | 1 | 4.75 | .25 | ||||||||||
| Totals or means, 4.09 | 1 | 3 | 3 | 43 | 207 | 217 | 75 | 23 | 11 | 4 | 3 | 590 | 3.94 | .17 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2¾ | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | 4¾ | ||||
| 4 | 4 | 17 | 23 | 19 | 2 | 65 | 3.67 | .31 | |||
| 4⅛ | 1 | 8 | 16 | 5 | 1 | 31 | 3.97 | .15 | |||
| 4¼ | 1 | 5 | 10 | 20 | 6 | 1 | 43 | 3.93 | .32 | ||
| 4⅜ | 1 | 5 | 3 | 1 | 10 | 3.85 | .52 | ||||
| 4½ | 1 | 14 | 15 | 7 | 3 | 40 | 3.98 | .52 | |||
| 4⅝ | 3 | 1 | 1 | 5 | 3.95 | .80 | |||||
| Totals or means, 4.22 | 1 | 4 | 25 | 63 | 74 | 21 | 5 | 1 | 194 | 3.88 | .34 |
| Generation. | No. of offspring. | Mean, parents. | Mean, offspring. | Standard deviation, parents. | Standard deviation, offspring. | Correlation, parents-offspring. | Absolute regression of offspring on parents. | Advance of parents. | Advance of offspring. |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 150 | 2.51 | 2.05 | .313 | .541 | .298 | .46 | ||
| 2 | 471 | 2.52 | 1.92 | .307 | .732 | .317 | .60 | .01 | -.13 |
| 3 | 341 | 2.73 | 2.51 | .285 | .531 | .331 | .22 | .21 | .59 |
| 4 | 444 | 3.09 | 2.73 | .215 | .468 | .066 | .36 | .36 | .22 |
| 5 | 610 | 3.33 | 2.90 | .240 | .505 | .160 | .43 | .24 | .17 |
| 6 | 861 | 3.52 | 3.11 | .209 | .490 | .180 | .41 | .19 | .21 |
| 7 | 1,077 | 3.56 | 3.20 | .212 | .555 | .215 | .36 | .04 | .09 |
| 8 | 1,408 | 3.75 | 3.48 | .246 | .439 | .099 | .27 | .19 | .28 |
| 9 | 1,322 | 3.78 | 3.54 | .112 | .346 | .210 | .24 | .03 | .06 |
| 10 | 776 | 3.88 | 3.73 | .112 | .362 | .116 | .15 | .10 | .19 |
| 11 | 697 | 3.98 | 3.78 | .113 | .289 | .233 | .20 | .10 | .05 |
| 12 | 590 | 4.09 | 3.94 | .176 | .302 | .161 | .15 | .11 | .16 |
| 13 | 194 | 4.22 | 3.88 | .433 | .270 | .132 | .34 | .13 | -.06 |
| Total | 8,941 |
| Generation. | Grade of parents; below, grade and number of their offspring. | Total number of offspring. | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 2⅛ | 2¼ | 2⅜ | 2½ | 2⅝ | 2¾ | 2⅞ | 3 | 3⅛ | 3¼ | 3⅜ | 3½ | 3⅝ | 3¾ | 3⅞ | 4 | 4⅛ | 4¼ | 4⅜ | 4½ | 4⅝ | 4¾ | ||
| 1 | 1.76 | 1.87 | 2.06 | 2.15 | 2.12 | 2.35 | ||||||||||||||||||
| 18 | 20 | 37 | 5 | 51 | 12 | |||||||||||||||||||
| 2 | 1.70 | 1.28 | 1.87 | 1.92 | 1.80 | 2.11 | 1.92 | 2.41 | 2.47 | 2.50 | ||||||||||||||
| 46 | 56 | 8 | 45 | 133 | 44 | 52 | 23 | 59 | 5 | |||||||||||||||
| 3 | 2.06 | 2.15 | 2.32 | 2.63 | 2.46 | 2.70 | 2.49 | 3.07 | 3.17 | 2.91 | ||||||||||||||
| 21 | 17 | 54 | 64 | 43 | 47 | 71 | 10 | 6 | 8 | |||||||||||||||
| 4 | 2.55 | 2.65 | 2.97 | 2.60 | 2.69 | 2.89 | 2.70 | 3.02 | 2.75 | |||||||||||||||
| 11 | 18 | 37 | 143 | 122 | 64 | 23 | 20 | 6 | ||||||||||||||||
| 5 | 3.00 | 2.87 | 2.81 | 2.81 | 2.81 | 2.94 | 2.87 | 3.08 | 3.07 | 3.35 | 3.36 | 3.00 | ||||||||||||
| 12 | 21 | 15 | 114 | 138 | 145 | 69 | 64 | 14 | 8 | 7 | 3 | |||||||||||||
| 6 | 3.25 | 2.84 | 3.10 | 2.96 | 3.10 | 3.16 | 3.22 | 3.26 | 3.41 | |||||||||||||||
| 2 | 28 | 143 | 123 | 212 | 181 | 78 | 80 | 14 | ||||||||||||||||
| 7 | 2.87 | 3.09 | 2.97 | 3.18 | 3.23 | 3.35 | 3.49 | 3.53 | 3.75 | |||||||||||||||
| 28 | 131 | 160 | 177 | 289 | 184 | 90 | 15 | 3 | 1,077 | |||||||||||||||
| 8 | 3.46 | 3.50 | 3.49 | 3.53 | 3.31 | 3.72 | 3.69 | 3.96 | ||||||||||||||||
| 59 | 484 | 469 | 238 | 64 | 60 | 22 | 8 | 1,408 | ||||||||||||||||
| 9 | 3.25 | 3.43 | 3.50 | 3.65 | 3.57 | 3.57 | 3.75 | 3.50 | ||||||||||||||||
| 4 | 244 | 591 | 424 | 45 | 7 | 4 | 3 | 1,322 | ||||||||||||||||
| 10 | 3.69 | 3.72 | 3.76 | 3.87 | 3.84 | |||||||||||||||||||
| 257 | 347 | 120 | 44 | 8 | 776 | |||||||||||||||||||
| 11 | 3.75 | 3.70 | 3.87 | 3.81 | 3.91 | 3.94 | ||||||||||||||||||
| 11 | 333 | 214 | 110 | 25 | 4 | 697 | ||||||||||||||||||
| 12 | 3.82 | 3.96 | 3.93 | 3.91 | 3.94 | 4.12 | 4.14 | 3.90 | 4.75 | |||||||||||||||
| 35 | 48 | 145 | 227 | 71 | 45 | 7 | 11 | 1 | 590 | |||||||||||||||
| 13 | 3.67 | 3.97 | 3.93 | 3.85 | 3.98 | 3.95 | ||||||||||||||||||
| 65 | 31 | 43 | 10 | 40 | 5 | 194 | ||||||||||||||||||
| 8,941 | ||||||||||||||||||||||||
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +¼ | 0 | ¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | ||||
| -1¼ | 1 | 2 | 3 | 2 | 8 | 1.34 | -.09 | ||||||
| 1⅜ | 1 | 1 | 3 | 12 | 8 | 3 | 1 | 1 | 1 | 31 | .86 | .51 | |
| 1½ | 1 | 1 | 1 | 1 | 1 | 1 | 6 | 1.37 | .13 | ||||
| 1⅞ | 1 | 1 | 2 | 2 | 3 | 1 | 10 | 1.05 | .82 | ||||
| Totals or means, 1.46 | 2 | 1 | 4 | 1 | 15 | 13 | 4 | 8 | 5 | 2 | 55 | 1.00 | .46 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +½ | 0 | -¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | ||||
| ¾ | 1 | 2 | 2 | 1 | 6 | 2 | 3 | 2 | 19 | 1.04 | -.29 | ||
| 1⅛ | 2 | 2 | 2 | 2 | 3 | 1 | 12 | 1.05 | .07 | ||||
| 1¼ | 4 | 6 | 3 | 3 | 1 | 17 | 1.18 | .07 | |||||
| 1⅜ | 1 | 1 | 1 | 2 | 5 | 1.45 | -.08 | ||||||
| 1½ | 1 | 10 | 13 | 1 | 8 | 4 | 37 | 1.11 | .39 | ||||
| 1⅝ | 1 | 1 | 1 | 3 | .67 | .95 | |||||||
| 1¾ | 1 | 1 | 2 | 14 | 4 | 3 | 1 | 1 | 27 | 1.09 | .66 | ||
| 1⅞ | 1 | 1 | 1 | 2 | 3 | 1 | 3 | 12 | 1.10 | .77 | |||
| Totals or means, 1.41 | 2 | 3 | 6 | 6 | 20 | 44 | 13 | 19 | 14 | 5 | 132 | 1.07 | .34 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | -¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | ||||
| -1⅛ | 2 | 1 | 1 | 1 | 5 | .85 | .27 | |||||
| 1¼ | 1 | 3 | 6 | 5 | 1 | 2 | 1 | 1 | 20 | 1.05 | .20 | |
| 1⅜ | 1 | 3 | 4 | 11 | 3 | 4 | 2 | 28 | 1.03 | .34 | ||
| 1½ | 1 | 1 | 12 | 1 | 5 | 5 | 3 | 28 | 1.31 | .19 | ||
| 1⅝ | 1 | 1 | 1 | 10 | 10 | 5 | 8 | 8 | 4 | 48 | 1.22 | .40 |
| 1¾ | 1 | 2 | 8 | 19 | 9 | 9 | 7 | 8 | 63 | 1.26 | .49 | |
| 1⅞ | 3 | 3 | 1.00 | .87 | ||||||||
| Totals or means, 1.56 | 1 | 4 | 12 | 30 | 61 | 19 | 29 | 23 | 16 | 195 | 1.18 | .38 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +½ | +¼ | 0 | -¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | ||||
| -1¼ | 1 | 1 | 1 | 1 | 4 | 1.56 | -.31 | ||||||||
| 1⅜ | 2 | 3 | 4 | 8 | 3 | 1 | 2 | 6 | 29 | 1.16 | .21 | ||||
| 1½ | 1 | 1 | 3 | 2 | 7 | 7 | 6 | 8 | 16 | 8 | 59 | 1.31 | .19 | ||
| 1⅝ | 1 | 4 | 10 | 4 | 8 | 7 | 6 | 40 | 1.36 | .26 | |||||
| 1¾ | 4 | 4 | 6 | 19 | 11 | 16 | 22 | 11 | 93 | 1.34 | .41 | ||||
| 1⅞ | 2 | 1 | 2 | 6 | 1 | 10 | 19 | 12 | 17 | 19 | 5 | 1 | 95 | 1.18 | .69 |
| 2 | 1 | 2 | 3 | 1 | 2 | 9 | 1.36 | .64 | |||||||
| Totals or means, 1.69 | 2 | 2 | 3 | 16 | 10 | 32 | 66 | 39 | 51 | 68 | 39 | 1 | 329 | 1.28 | .41 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | -¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | ||||
| ⅞ | 2 | 1 | 4 | 5 | 3 | 4 | 1 | 20 | 1.09 | -.22 | ||||
| 1 | 2 | 1 | 1 | 4 | .99 | .01 | ||||||||
| 1⅜ | 4 | 1 | 1 | 6 | 2 | 10 | 14 | 11 | 2 | 51 | 1.50 | -.13 | ||
| 1½ | 1 | 3 | 9 | 9 | 11 | 7 | 7 | 5 | 1 | 53 | 1.25 | .25 | ||
| 1⅝ | 1 | 1 | 5 | 12 | 4 | 15 | 12 | 4 | 54 | 1.35 | .27 | |||
| 1¾ | 9 | 14 | 29 | 50 | 30 | 50 | 55 | 24 | 1 | 262 | 1.30 | .45 | ||
| 1⅞ | 4 | 11 | 12 | 29 | 50 | 31 | 6 | 143 | 1.64 | .23 | ||||
| 2 | 1 | 2 | 11 | 14 | 5 | 19 | 28 | 25 | 4 | 109 | 1.52 | .48 | ||
| 2⅛ | 1 | 2 | 2 | 5 | 1.80 | .32 | ||||||||
| Totals or means, 1.73 | 3 | 18 | 21 | 63 | 108 | 64 | 134 | 172 | 104 | 13 | 1 | 701 | 1.41 | .32 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | -¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | ||||
| -1⅛ | 1 | 1 | 2 | 1 | 5 | .85 | .27 | |||||||
| 1⅜ | 1 | 1 | 1 | 1 | 4 | 1.44 | -.07 | |||||||
| 1½ | 1 | 3 | 4 | 4 | 4 | 6 | 2 | 24 | 1.34 | .16 | ||||
| 1⅝ | 1 | 1 | 11 | 16 | 3 | 17 | 28 | 17 | 94 | 1.45 | .17 | |||
| 1¾ | 1 | 1 | 8 | 14 | 39 | 20 | 39 | 77 | 43 | 2 | 244 | 1.49 | .26 | |
| 1⅞ | 1 | 9 | 24 | 61 | 40 | 71 | 156 | 127 | 12 | 1 | 502 | 1.59 | .28 | |
| 2 | 1 | 3 | 5 | 12 | 32 | 22 | 37 | 89 | 76 | 5 | 1 | 283 | 1.58 | .42 |
| 2⅛ | 2 | 4 | 4 | 5 | 35 | 31 | 4 | 85 | 1.52 | .60 | ||||
| 2½ | 2 | 5 | 3 | 1 | 11 | 1.82 | .68 | |||||||
| Totals or means, 1.86 | 3 | 6 | 25 | 66 | 159 | 94 | 177 | 396 | 300 | 22 | 4 | 1252 | 1.56 | .30 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | ¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | ||||
| 1⅝ | 1 | 1 | 2 | 1 | 5 | 1.55 | .07 | ||||||||
| 1¾ | 1 | 2 | 1 | 1 | 6 | 11 | 9 | 1 | 32 | 1.67 | .08 | ||||
| 1⅞ | 2 | 1 | 6 | 4 | 35 | 20 | 47 | 99 | 106 | 6 | 3 | 1 | 330 | 1.65 | .22 |
| 2 | 8 | 14 | 51 | 60 | 119 | 368 | 324 | 22 | 3 | 969 | 1.72 | .28 | |||
| 2⅛ | 1 | 2 | 3 | 14 | 10 | 30 | 92 | 104 | 4 | 260 | 1.74 | .38 | |||
| 2¼ | 5 | 10 | 19 | 6 | 1 | 41 | 1.93 | .32 | |||||||
| 2⅜ | 2 | 2 | 4 | 1.63 | .74 | ||||||||||
| 2½ | 1 | 4 | 10 | 15 | 1.88 | .62 | |||||||||
| 2⅝ | 1 | 3 | 8 | 5 | 1 | 18 | 2.28 | .09 | |||||||
| 2¾ | 3 | 3 | 6 | 1.87 | .88 | ||||||||||
| Totals or means, 2.01 | 2 | 2 | 17 | 24 | 101 | 92 | 211 | 594 | 584 | 43 | 8 | 2 | 1680 | 1.73 | .28 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | ||||
| 1⅞ | 1 | 9 | 8 | 1 | 19 | 1.84 | .03 | |||||||
| 2 | 6 | 8 | 38 | 22 | 122 | 550 | 423 | 21 | 9 | 3 | 1202 | 1.81 | .19 | |
| 2⅛ | 1 | 3 | 12 | 7 | 31 | 118 | 187 | 32 | 2 | 3 | 396 | 1.86 | .26 | |
| 2¼ | 1 | 2 | 19 | 20 | 3 | 45 | 1.87 | .38 | ||||||
| 2⅜ | 1 | 1 | 11 | 17 | 30 | 1.87 | .50 | |||||||
| 2½ | 3 | 6 | 15 | 4 | 28 | 1.93 | .57 | |||||||
| 2⅝ | 3 | 2 | 1 | 6 | 1.67 | .95 | ||||||||
| Totals or means, 2.05 | 1 | 6 | 11 | 52 | 30 | 162 | 715 | 671 | 61 | 11 | 6 | 1726 | 1.80 | .25 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | ||||
| 2 | 1 | 13 | 4 | 36 | 268 | 420 | 55 | 10 | 4 | 811 | 1.90 | .10 | |
| 2⅛ | 1 | 1 | 25 | 110 | 218 | 40 | 7 | 1 | 403 | 1.93 | .19 | ||
| 2¼ | 1 | 1 | 7 | 43 | 77 | 12 | 6 | 1 | 148 | 1.93 | .32 | ||
| 2⅜ | 1 | 1 | 7 | 61 | 89 | 12 | 4 | 175 | 1.91 | .46 | |||
| 2½ | 1 | 2 | 6 | 23 | 14 | 5 | 2 | 53 | 2.07 | .43 | |||
| 2⅝ | 1 | 1 | 2.00 | .62 | |||||||||
| Totals or means, 2.11 | 1 | 1 | 16 | 7 | 77 | 488 | 828 | 133 | 32 | 8 | 1591 | 1.92 | .19 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | ||||
| 2 | 1 | 2 | 13 | 120 | 287 | 43 | 7 | 2 | 2 | 1 | 478 | 1.96 | .04 |
| 2⅛ | 2 | 1 | 15 | 100 | 251 | 78 | 22 | 4 | 1 | 474 | 2.00 | .12 | |
| 2¼ | 5 | 45 | 92 | 58 | 13 | 4 | 217 | 2.05 | .20 | ||||
| 2⅜ | 1 | 4 | 16 | 60 | 29 | 6 | 3 | 119 | 2.05 | .32 | |||
| 2½ | 9 | 27 | 19 | 10 | 2 | 67 | 2.13 | .37 | |||||
| 2⅝ | 1 | 5 | 17 | 17 | 8 | 1 | 49 | 2.15 | .47 | ||||
| 2¾ | 1 | 1 | 8 | 10 | 6 | 1 | 27 | 1.95 | .80 | ||||
| 2⅞ | 1 | 1 | 2 | 3 | 6 | 5 | 2 | 20 | 2.19 | .68 | |||
| Totals or means, 2.18 | 3 | 6 | 40 | 305 | 747 | 256 | 72 | 17 | 3 | 2 | 1451 | 2.01 | .17 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | ||||
| 2 | 7 | 26 | 14 | 3 | 1 | 51 | 2.08 | .08 | ||||||
| 2⅛ | 4 | 20 | 75 | 45 | 26 | 12 | 1 | 183 | 2.15 | -.03 | ||||
| 2¼ | 1 | 12 | 49 | 111 | 76 | 50 | 13 | 6 | 318 | 2.13 | .12 | |||
| 2⅜ | 2 | 1 | 4 | 24 | 112 | 66 | 33 | 20 | 4 | 2 | 268 | 2.16 | .21 | |
| 2½ | 1 | 4 | 32 | 21 | 16 | 7 | 81 | 2.21 | .29 | |||||
| 2⅝ | 1 | 3 | 18 | 15 | 11 | 4 | 52 | 2.20 | .42 | |||||
| 2¾ | 1 | 4 | 9 | 4 | 3 | 1 | 1 | 2 | 1 | 26 | 2.26 | .49 | ||
| 2⅞ | 2 | 2 | 1 | 5 | 1.95 | .92 | ||||||||
| Totals or means, 2.30 | 3 | 2 | 22 | 113 | 385 | 242 | 142 | 57 | 12 | 5 | 1 | 984 | 2.15 | .15 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | ||||
| 2 | 2 | 7 | 1 | 10 | 1.98 | .02 | |||||||
| 2⅛ | 1 | 6 | 65 | 26 | 15 | 4 | 1 | 118 | 2.14 | -.02 | |||
| 2¼ | 1 | 2 | 16 | 67 | 45 | 29 | 6 | 155 | 2.15 | .10 | |||
| 2⅜ | 1 | 21 | 116 | 64 | 24 | 3 | 1 | 230 | 2.11 | .26 | |||
| 2½ | 1 | 12 | 81 | 75 | 53 | 17 | 3 | 242 | 2.24 | .26 | |||
| 2⅝ | 5 | 38 | 36 | 37 | 12 | 8 | 1 | 137 | 2.32 | .30 | |||
| 2¾ | 8 | 7 | 14 | 10 | 3 | 42 | 2.45 | .30 | |||||
| 2⅞ | 1 | 7 | 9 | 14 | 15 | 7 | 53 | 2.51 | .36 | ||||
| 3 | 5 | 5 | 5 | 10 | 5 | 2 | 1 | 35 | 2.65 | .35 | |||
| Totals or means, 2.44 | 1 | 5 | 63 | 394 | 268 | 191 | 83 | 27 | 3 | 2 | 1037 | 2.23 | .21 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | ||||
| 2¼ | 3 | 30 | 12 | 17 | 4 | 1 | 67 | 2.22 | .03 | ||
| 2⅜ | 4 | 40 | 50 | 46 | 24 | 5 | 1 | 170 | 2.35 | .02 | |
| 2½ | 4 | 32 | 34 | 47 | 26 | 8 | 2 | 1 | 154 | 2.40 | .10 |
| 2⅝ | 2 | 11 | 21 | 41 | 28 | 3 | 2 | 108 | 2.47 | .15 | |
| 2¾ | 1 | 5 | 2 | 9 | 7 | 6 | 1 | 31 | 2.56 | .19 | |
| 2⅞ | 4 | 7 | 8 | 6 | 1 | 26 | 2.43 | .44 | |||
| 3 | 3 | 1 | 5 | 5 | 1 | 15 | 2.50 | .50 | |||
| Totals or means, 2.50 | 14 | 125 | 127 | 173 | 100 | 25 | 6 | 1 | 571 | 2.39 | .11 |
| Generation. | No. of offspring. | Mean, parents. | Mean, offspring. | Standard deviation, parents. | Standard deviation, offspring. | Correlation, parents-offspring. | Absolute regression of offspring on parents. | Advance of parents. | Advance of offspring. |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 5 | 1.46 | 1.00 | .208 | .515 | .46 | |||
| 2 | 13 | 1.41 | 1.07 | .342 | .493 | -.033 | .34 | -.05 | .07 |
| 3 | 19 | 1.56 | 1.18 | .196 | .484 | .206 | .38 | .15 | .11 |
| 4 | 32 | 1.69 | 1.28 | .190 | .460 | .020 | .41 | .13 | .10 |
| 5 | 70 | 1.73 | 1.41 | .233 | .500 | .184 | .32 | .04 | .13 |
| 6 | 1,25 | 1.86 | 1.56 | .185 | .488 | .164 | .30 | .13 | .15 |
| 7 | 1,68 | 2.01 | 1.73 | .132 | .352 | .143 | .28 | .15 | .17 |
| 8 | 1,72 | 2.05 | 1.80 | .107 | .283 | .094 | .25 | .04 | .07 |
| 9 | 1,59 | 2.11 | 1.92 | .184 | .285 | .059 | .19 | .06 | .12 |
| 10 | 1,45 | 2.18 | 2.01 | .255 | .242 | .158 | .17 | .07 | .09 |
| 11 | 98 | 2.30 | 2.15 | .229 | .349 | .081 | .15 | .12 | .14 |
| 12 | 1,03 | 2.44 | 2.23 | .310 | .372 | .406 | .07 | .14 | .08 |
| 13 | 57 | 2.50 | 2.39 | .177 | .317 | .235 | .11 | .06 | .16 |
| Total | 11,704 |
| Generation. | Grade of parents; below, grade and number of their offspring. | Total number of offspring. | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1¼ | 1⅜ | 1½ | 1⅝ | 1¾ | 1⅞ | 2 | 2⅛ | 2¼ | 2⅜ | 2½ | 2⅝ | 2¾ | 2⅞ | 3 | ||
| 1 | 1.34 | .85 | 1.37 | 1.05 | ||||||||||||
| 8 | 31 | 6 | 10 | 55 | ||||||||||||
| 2 | 1.17 | 1.45 | 1.11 | 0.67 | 1.09 | 1.10 | ||||||||||
| 17 | 5 | 37 | 3 | 27 | 12 | 132 | ||||||||||
| 3 | 1.05 | 1.04 | 1.31 | 1.22 | 1.26 | 1.96 | ||||||||||
| 20 | 28 | 28 | 48 | 63 | 3 | 195 | ||||||||||
| 4 | 1.56 | 1.16 | 1.31 | 1.36 | 1.34 | 1.18 | 1.36 | |||||||||
| 4 | 29 | 59 | 40 | 93 | 95 | 9 | 329 | |||||||||
| 5 | 1.50 | 1.25 | 1.35 | 1.30 | 1.64 | 1.52 | 1.80 | |||||||||
| 51 | 53 | 54 | 262 | 143 | 109 | 5 | 701 | |||||||||
| 6 | 1.34 | 1.46 | 1.49 | 1.59 | 1.58 | 1.52 | 1.82 | |||||||||
| 24 | 94 | 244 | 502 | 283 | 85 | 11 | 1,252 | |||||||||
| 7 | 1.55 | 1.67 | 1.65 | 1.72 | 1.74 | 1.93 | 1.62 | 1.88 | 2.28 | 1.87 | ||||||
| 5 | 32 | 330 | 969 | 260 | 41 | 4 | 15 | 18 | 6 | 1,680 | ||||||
| 8 | 1.84 | 1.81 | 1.86 | 1.87 | 1.87 | 1.92 | 1.67 | |||||||||
| 19 | 1170 | 377 | 36 | 30 | 17 | 6 | 1,726 | |||||||||
| 9 | 1.90 | 1.93 | 1.93 | 1.91 | 2.07 | |||||||||||
| 811 | 403 | 148 | 175 | 53 | 1,591 | |||||||||||
| 10 | 1.96 | 2.00 | 2.04 | 2.05 | 2.13 | 2.15 | 1.95 | 2.18 | ||||||||
| 478 | 474 | 217 | 119 | 67 | 49 | 27 | 20 | 1,451 | ||||||||
| 11 | 2.08 | 2.15 | 2.13 | 2.16 | 2.21 | 2.20 | 2.26 | 1.95 | ||||||||
| 51 | 183 | 318 | 268 | 81 | 52 | 26 | 5 | 984 | ||||||||
| 12 | 1.98 | 2.14 | 2.15 | 2.11 | 2.24 | 2.32 | 2.45 | 2.51 | 2.65 | |||||||
| 10 | 118 | 166 | 230 | 242 | 137 | 42 | 53 | 35 | 1,037 | |||||||
| 13 | 2.22 | 2.35 | 2.40 | 2.47 | 2.55 | 2.43 | 2.50 | |||||||||
| 67 | 170 | 154 | 108 | 31 | 26 | 15 | 571 | |||||||||
| 11,704 | ||||||||||||||||
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | ¼ | ½ | ¾ | -1 | -1¼ | -1½ | -1¾ | -2 | ||||
| ⅜ | 2 | 1 | 3 | 4 | 8 | 8 | 26 | 1.08 | -.60 | |||
| ⅝ | 2 | 2 | 3 | 4 | 10 | 14 | 17 | 20 | 4 | 76 | 1.33 | -.71 |
| ¾ | 1 | 1 | 2 | 1 | 5 | 1.30 | -.65 | |||||
| ⅞ | 2 | 2 | 1 | 4 | 2 | 11 | 1.41 | -.63 | ||||
| Totals or means, .60 | 2 | 4 | 6 | 5 | 16 | 18 | 28 | 33 | 6 | 118 | 1.28 | -.68 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | ½ | ¾ | -1 | -1¼ | -1½ | ||||
| ½ | 1 | 3 | 4 | 5 | 2 | 4 | 19 | .95 | -.45 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | ¼ | ½ | ¾ | -1 | -1¼ | -1½ | -1¾ | -2 | ||||
| ¼ | 5 | 2 | 2 | 3 | 1 | 13 | .63 | -.38 | ||||
| ⅜ | 2 | 2 | 3 | 5 | 4 | 2 | 3 | 21 | .86 | -.49 | ||
| -1⅛ | 1 | 2 | 3 | 4 | 3 | 13 | 1.61 | -.49 | ||||
| -1¼ | 1 | 2 | 4 | 9 | 8 | 5 | 2 | 31 | 1.35 | -.10 | ||
| Totals or means, .83 | 2 | 7 | 6 | 9 | 12 | 11 | 14 | 12 | 5 | 78 | 1.14 | -.31 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | ¼ | ½ | ¾ | -1 | -1¼ | -1½ | -1¾ | -2 | ||||
| ¼ | 1 | 2 | 1 | 1 | 4 | 2 | 3 | 2 | 16 | 1.34 | -1.09 | |
| ½ | 1 | 1 | 1 | 2 | 5 | .70 | -.20 | |||||
| ⅝ | 1 | 1 | 1 | 3 | .83 | -.21 | ||||||
| ¾ | 1 | 1 | 1 | 2 | 1 | 6 | .83 | -.08 | ||||
| ⅞ | 1 | 2 | 2 | 2 | 2 | 1 | 10 | 1.17 | -.30 | |||
| -1 | 2 | 1 | 3 | 3 | 1 | 10 | 1.42 | -.42 | ||||
| Totals or means, .63 | 3 | 2 | 6 | 4 | 7 | 10 | 8 | 6 | 4 | 50 | 1.17 | -.54 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | ¼ | ½ | ¾ | -1¼ | -1½ | ||||
| ¼ | 2 | 2 | 4 | 1.00 | -.75 | ||||
| ¾ | 1 | 3 | 4 | 1.12 | -.37 | ||||
| ⅞ | 2 | 2 | 1 | 5 | .25 | +.62 | |||
| Totals or means, .65 | 3 | 2 | 1 | 2 | 2 | 3 | 13 | .75 | -.10 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +1¼ | +1 | +¾ | +½ | +¼ | 0 | ¼ | ½ | ¾ | -1 | -1¼ | -1½ | -1¾ | ||||
| ⅛ | 1 | 1 | 3 | 2 | 1 | 2 | 2 | 12 | .29 | -.17 | ||||||
| ¼ | 2 | 1 | 1 | 4 | 1 | 4 | 13 | .35 | -.10 | |||||||
| ⅜ | 1 | 2 | 1 | 1 | 1 | 6 | .87 | -.50 | ||||||||
| ½ | 1 | 1 | 2 | 1 | 5 | .25 | +.25 | |||||||||
| Totals or means, .26 | 1 | 1 | 1 | 2 | 1 | 4 | 7 | 2 | 5 | 8 | 2 | 1 | 1 | 36 | .39 | -.13 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ½ | ¾ | -1 | -1¼ | -1½ | -1¾ | -2 | ||||
| ¾ | 1 | 3 | 3 | 7 | 8 | 3 | 25 | |||
| ⅞ | 2 | 2 | 2 | 2 | 8 | |||||
| Totals or means, .78 | 2 | 3 | 5 | 3 | 7 | 10 | 3 | 33 | 1.15 | -.37 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | |||||
|---|---|---|---|---|---|---|---|---|---|
| ¾ | -1 | -1¼ | -1½ | -1¾ | -2 | ||||
| ½ | 1 | 3 | 6 | 3 | 13 | 1.69 | -1.19 | ||
| ⅝ | 1 | 9 | 3 | 13 | 1.21 | - .59 | |||
| ⅞ | 2 | 2 | 2.00 | - .13 | |||||
| -1 | 1 | 2 | 4 | 4 | 2 | 13 | 1.56 | - .56 | |
| Totals or means, .72 | 1 | 2 | 2 | 7 | 19 | 10 | 41 | 1.51 | - .79 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||
|---|---|---|---|---|---|---|---|---|
| ¼ | -1¾ | -2 | -2¼ | -2½ | ||||
| -1⅝ | 1 | 3 | 7 | 3 | 2 | 16 | 1.95 | -.33 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +½ | +¾ | +2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | ||||
| 2 | 1 | 1 | 3 | 3 | 2 | 2 | 3 | 1 | 1 | 17 | 2.36 | -.36 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Regression. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | ||||
| 2⅝ | 2 | 3 | 2 | 5 | 5 | 1 | 1 | 22 | ||||
| 2¾ | 2 | 4 | 1 | 8 | 2 | 17 | ||||||
| 3 | 4 | 4 | 1 | 9 | ||||||||
| 3¼ | 1 | 3 | 1 | 5 | ||||||||
| Totals or means, 2.79 | 2 | 3 | 4 | 10 | 6 | 12 | 10 | 5 | 1 | 53 | 3.32 | -.53 |
| Grade of hooded grandparents. | Grade of extracted hooded grandchildren. | Total. | Mean. | Mean, uncrossed race. | Standard deviation. | Standard deviation, uncrossed race. | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -2 | -1¾ | -1½ | -1¼ | -1 | -¾ | -½ | -¼ | 0 | +¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | ||||||
| ♀ -1⅞, gen. 2½, minus series | 2 | 4 | 6 | 5 | 6 | 13 | 2 | 1 | 3 | 4 | 5 | 3 | 4 | 2 | 1 | 1 | 62 | + .31 | -1.20 | .98 | .49 | ||||||||
| ♀♀ -2, gen. 6, minus series | 1 | 1 | 1 | 1 | 2 | 6 | 6 | 7 | 2 | 3 | 6 | 4 | 2 | 4 | 1 | 1 | 48 | + .25 | -1.59 | .97 | .44 | ||||||||
| ♀♀ -2 and -2¼, gen. 10, minus series | 1 | 3 | 3 | 1 | 10 | 3 | 9 | 8 | 14 | 4 | 7 | 8 | 1 | 4 | 3 | 5 | 2 | 4 | 1 | 91 | + .24 | -2.05 | 1.18 | .24 | |||||
| ♀♀ +3, gen. 3, plus series | 1 | 5 | 3 | 2 | 3 | 4 | 2 | 1 | 21 | +2.56 | +2.60 | .50 | .53 | ||||||||||||||||
| ♀♀ +3¼, gen. 5 and +3½, gen. 6, plus series | 1 | 2 | 1 | 1 | 1 | 6 | 9 | 8 | 8 | 1 | 38 | +2.97 | +3.14 | .52 | .49 | ||||||||||||||
| ♀ +4¼, gen. 10, plus series | 1 | 3 | 4 | 2 | 3 | 3 | 16 | +3.15 | +3.84 | .28 | .36 | ||||||||||||||||||
| Grade of hooded grandparents. | Grade of extracted hooded grandchildren. | Total. | Mean. | Mean, same grade and generation, uncrossed race. | Standard deviation. | Standard deviation, uncrossed race. | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -2 | -1¾ | -1½ | -1¼ | -1 | -¾ | -½ | -¼ | 0 | +¼ | +½ | +¾ | +1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | ||||||
| -1½, gen. 3½ | 1 | 5 | 3 | 6 | 12 | 26 | 9 | 9 | 10 | 1 | 3 | 2 | 2 | 1 | 90 | - .62 | -1.31 | .61 | .48 | ||||||||
| -1⅞, gen. 4 | 6 | 3 | 4 | 5 | 9 | 14 | 8 | 1 | 1 | 1 | 1 | 53 | - .73 | -1.18 | .60 | .46 | |||||||||||
| +2, gen. 7½ | 6 | 6 | 12 | 11 | 8 | 3 | 4 | 4 | 6 | 1 | 3 | 1 | 1 | 66 | - .94 | -1.75 | .84 | .35 | |||||||||
| +2¼, gen. 2 | 3 | 4 | 5 | 4 | 16 | 7 | 16 | 22 | 25 | 29 | 24 | 13 | 33 | 14 | 8 | 8 | 6 | 3 | 239 | +1.27 | +1.80 | .90 | .75 | ||||
| +3, gen. 3 | 2 | 1 | 1 | 3 | 3 | 4 | 2 | 2 | 1 | 2 | 1 | 1 | 23 | + .95 | +2.50 | .87 | .53 | ||||||||||
| Grade of (F₂) parents. | Grade of (F₃) offspring. | Totals. | Means. | Regression. | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -1 | -¾ | -½ | -¼ | 0 | +½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | ||||
| ¾ | 2 | 1 | 4 | 3 | 4 | 1 | 2 | 1 | 1 | 19 | + .04 | .71 | |||||||||
| 2½ | 1 | 1 | 1 | 3 | 2 | 2 | 1 | 5 | 3 | 6 | 4 | 3 | 1 | 1 | 34 | +2.06 | .44 | ||||
| Grade of (F₃) parents. | Grade of (F₄) offspring. | Totals. | Means. | Regression. | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -¼ | 0 | +¼ | +½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | ||||
| 2¼ | 2 | 1 | 1 | 3 | 6 | 6 | 9 | 16 | 25 | 14 | 10 | 2 | 1 | 96 | 1.78 | .47 | |||
| 2⅜ | 2 | 1 | 1 | 1 | 6 | 3 | 8 | 3 | 9 | 4 | 4 | 1 | 1 | 44 | 1.67 | .70 | |||
| 2⅞ | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 8 | .91 | 1.96 | |||||||||
| 3 | 1 | 1 | 3 | 3 | 5 | 9 | 5 | 7 | 3 | 1 | 1 | 39 | 2.02 | .98 | |||||
| 3⅛ | 1 | 1 | 3 | 1 | 2 | 5 | 4 | 1 | 18 | 2.82 | .30 | ||||||||
| Totals or means, 2.52 | 3 | 3 | 1 | 5 | 5 | 16 | 10 | 22 | 26 | 47 | 20 | 23 | 14 | 7 | 1 | 2 | 205 | 1.86 | .66 |
| Grade of (F₄) parents. | Grade of (F₅) offspring. | Totals. | Means. | Regression. | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | +½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | ||||
| 2 | 1 | 1 | 3 | 3 | 2 | 12 | 15 | 10 | 2 | 3 | 1 | 53 | ||||
| 2⅛ | 1 | 1 | 1 | 4 | 1 | 4 | 1 | 13 | ||||||||
| 2¼ | 1 | 3 | 4 | 1 | 2 | 2 | 13 | |||||||||
| 2⅜ | 1 | 2 | 3 | 2 | 4 | 2 | 14 | |||||||||
| 2⅝ | 1 | 1 | 2 | 1 | 2 | 7 | ||||||||||
| 2⅞ | 2 | 1 | 1 | 1 | 5 | |||||||||||
| 3 | 1 | 1 | 1 | 7 | 4 | 14 | ||||||||||
| Totals or means, 2.27 | 2 | 1 | 3 | 6 | 3 | 6 | 17 | 28 | 16 | 8 | 21 | 7 | 1 | 119 | 2.06 | .21 |
| Grade of (F₅) parents. | Grade of (F₆) offspring. | Totals. | Means. | Regression. | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | ||||
| 2⅜ | 1 | 1 | 1 | 4 | 8 | 13 | 4 | 4 | 7 | 3 | 2 | 48 | 2.14 | .23 | |||
| 2½ | 3 | 2 | 2 | 4 | 3 | 3 | 5 | 4 | 26 | 2.24 | .26 | ||||||
| 2⅝ | 1 | 1 | 1 | 2 | 1 | 2 | 3 | 3 | 4 | 1 | 1 | 20 | 2.26 | .36 | |||
| 2¾ | 1 | 1 | 4 | 2 | 5 | 3 | 8 | 2 | 5 | 31 | 2.35 | .40 | |||||
| 2⅞ | 1 | 4 | 1 | 9 | 7 | 11 | 5 | 2 | 40 | 2.74 | .13 | ||||||
| 3⅛ | 1 | 4 | 2 | 4 | 7 | 6 | 2 | 1 | 2 | 29 | 2.73 | .39 | |||||
| Totals or means, 2.69 | 2 | 1 | 2 | 6 | 13 | 14 | 32 | 13 | 23 | 37 | 30 | 15 | 4 | 2 | 194 | 2.41 | .28 |
| Grade of (F₆) parents. | Grade of (F₇) offspring. | Totals. | Means. | Regression. | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ¾ | 1 | 1½ | 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | ||||
| 2⅝ | 4 | 4 | 12 | 2 | 9 | 9 | 12 | 1 | 53 | 2.41 | .21 | ||||
| 2¾ | 3 | 1 | 2 | 6 | 5 | 4 | 4 | 3 | 28 | 2.14 | .61 | ||||
| 2⅞ | 1 | 1 | 1 | 2 | 3 | 1 | 1 | 10 | 2.32 | .55 | |||||
| 3 | 1 | 3 | 1 | 3 | 1 | 3 | 12 | 2.93 | .07 | ||||||
| 3⅛ | 1 | 3 | 2 | 3 | 1 | 4 | 3 | 5 | 2 | 2 | 1 | 27 | 2.59 | .53 | |
| 3¼ | 3 | 1 | 3 | 3 | 1 | 3 | 14 | 2.63 | .62 | ||||||
| 3⅜ | 1 | 2 | 2 | 2 | 2 | 1 | 10 | 2.85 | .52 | ||||||
| Totals or means, 2.80 | 3 | 2 | 8 | 9 | 26 | 12 | 28 | 23 | 26 | 10 | 6 | 1 | 154 | 2.46 | .34 |
| Grade of (F₇) parents. | Grade of (F₈) offspring. | Totals. | Means. | Regression. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1¾ | 2 | 2¼ | 2½ | 2¾ | 3 | 3¼ | 3½ | 3¾ | ||||
| 2¾ | 1 | 1 | 2 | 2.75 | 0 | |||||||
| 3 | 1 | 1 | 3 | 5 | 8 | 5 | 1 | 24 | 2.74 | .01 | ||
| 3⅛ | 1 | 2 | 3 | 3 | 5 | 3 | 1 | 1 | 19 | 2.60 | .52 | |
| Totals or means, 3.08 | 2 | 3 | 6 | 9 | 13 | 4 | 6 | 1 | 1 | 45 | 2.67 | .41 |
| Grade of parents. | Grade of offspring. | Totals. | Means. | Standard deviation. | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -2 | -1¾ | -1½ | -1¼ | -1 | ¾ | ½ | ¼ | 0 | +¼ | +½ | +¾ | +1 | +1¼ | +1½ | +1¾ | +2 | +2¼ | +2½ | +2¾ | +3½ | ||||
| Series 1, F₁ generation; ♀♀ -2, gen. 6; ♂ ♂ +3½ or 3¾, gen. 5 | 2 | 2 | 1 | 8 | 10 | 13 | 25 | 2 | 8 | 11 | 6 | 3 | 1 | 1 | 93 | + .06 | .71 | |||||||
| Series 1, F₂ generation, -1 | 1 | 1 | 4 | 1 | 1 | 3 | 1 | 1 | 13 | - .57 | .66 | |||||||||||||
| ½ | 1 | 2 | 1 | 3 | 3 | 2 | 1 | 1 | 1 | 1 | 16 | - .37 | .80 | |||||||||||
| 0 | 1 | 1 | 4 | 2 | 4 | 6 | 1 | 1 | 1 | 3 | 1 | 2 | 1 | 1 | 29 | + .17 | .95 | |||||||
| +⅛ | 1 | 2 | 1 | 3 | 3 | 1 | 5 | 1 | 4 | 3 | 2 | 3 | 1 | 30 | + .03 | .97 | ||||||||
| +¼ | 2 | 1 | 2 | 7 | 3 | 1 | 2 | 4 | 1 | 2 | 2 | 6 | 1 | 3 | 3 | 1 | 41 | + .10 | 1.08 | |||||
| +½ | 1 | 1 | 1 | 4 | 2 | 4 | 1 | 1 | 1 | 1 | 1 | 18 | + .06 | .91 | ||||||||||
| +¾ | 1 | 2 | 3 | 1 | 2 | 1 | 3 | 2 | 1 | 1 | 1 | 18 | + .25 | .90 | ||||||||||
| +1 | 1 | 7 | 3 | 6 | 13 | 4 | 8 | 5 | 5 | 4 | 4 | 3 | 2 | 2 | 6 | 73 | + .60 | .98 | ||||||
| +1⅛ | 1 | 1 | 1 | 2 | 1 | 3 | 2 | 1 | 1 | 13 | + .61 | .83 | ||||||||||||
| +1¼ | 1 | 1 | 1 | 5 | 7 | 1 | 8 | 1 | 5 | 6 | 4 | 2 | 4 | 3 | 1 | 3 | 53 | + .38 | 1.02 | |||||
| +1½ | 1 | 1 | 0 | |||||||||||||||||||||
| Series 1, totals or means, F₂ generation | 1 | 4 | 9 | 7 | 22 | 24 | 26 | 20 | 49 | 10 | 24 | 24 | 23 | 10 | 20 | 13 | 7 | 5 | 7 | 305 | + .24 | 1.01 | ||
| Series 2, F₁ generation; ♀ ♀ +3¾, gen. 10; ♂ -3¼, gen. 10 | 1 | 1 | 1 | 2 | 4 | 2 | 1 | 1 | 1 | 14 | +1.00 | .60 | ||||||||||||
| Series 2, F₂ generation, +½ | 1 | 1 | 1 | 3 | +1.31 | |||||||||||||||||||
| +⅝ | 2 | 1 | 3 | 3 | 3 | 1 | 1 | 1 | 1 | 16 | +.40 | .96 | ||||||||||||
| +1 | 2 | 2 | 1 | 1 | 1 | 7 | +.82 | |||||||||||||||||
| +1⅛ | 1 | 1 | 2 | 1 | 3 | 4 | 2 | 1 | 4 | 1 | 1 | 1 | 22 | +1.03 | .97 | |||||||||
| +1¼ | 2 | 1 | 1 | 1 | 4 | 5 | 3 | 2 | 4 | 1 | 1 | 25 | + .56 | .70 | ||||||||||
| Series 2, totals or means, F₂ generation | 2 | 4 | 3 | 8 | 8 | 12 | 9 | 6 | 5 | 3 | 5 | 2 | 2 | 1 | 2 | 1 | 73 | +.72 | .87 | |||||
| Grade of mother. | Grade of offspring. | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lower group. | Upper group. | |||||||||||||||||
| +3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | Totals. | Means. | Standard deviation. | 5 | 5¼ | 5½ | 5¾ | 6 | Totals. | Means. | Standard deviation. | |
| +3¾ | 1 | 4 | 11 | 1 | 1 | 18 | 3.69 | 1 | 3 | 9 | 4 | 1 | 18 | 5.51 | ||||
| 3⅞ | 3 | 7 | 3 | 13 | 3.75 | 1 | 4 | 5 | 3 | 13 | 5.44 | |||||||
| 4 | 1 | 4 | 9 | 4 | 1 | 1 | 20 | 3.78 | 2 | 7 | 5 | 4 | 18 | 5.40 | ||||
| 4⅛ | 3 | 1 | 1 | 5 | 3.90 | 2 | 4 | 1 | 7 | 5.46 | ||||||||
| 4½ | 1 | 1 | 2 | 3.62 | ||||||||||||||
| Totals or means | 1 | 1 | 12 | 31 | 9 | 3 | 1 | 58 | 3.73 | .24 | 4 | 16 | 23 | 12 | 1 | 56 | 5.45 | .23 |
| Mutant ♀ 5153, +5¾ | 3 | 3 | 6 | 3.87 | 6 | 4 | 10 | 5.60 | ||||||||||
| Grade of mother. | Grade of offspring, lower group. | Grade of offspring, upper group. | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -1 | ¾ | ½ | ¼ | 0 | +¼ | +½ | +¾ | +1 | +1¼ | +1½ | +1¾ | +2¼ | Totals. | Means. | Standard deviation. | +4 | +4¼ | +4½ | +4¾ | +5 | Totals. | Means. | Standard deviation. | |
| -2 | 1 | 1 | 1 | 3 | 1 | 2 | 1 | 1 | 1 | 1 | 13 | +.40 | 2 | 5 | 3 | 2 | 12 | 4.46 | ||||||
| -2¼ | 1 | 1 | 1 | 2 | 2 | 2 | 3 | 1 | 1 | 14 | +.70 | 2 | 4 | 3 | 9 | 4.53 | ||||||||
| -2½ | 2 | 1 | 1 | 4 | +.31 | 2 | 3 | 1 | 6 | 4.21 | ||||||||||||||
| -2¾ | 1 | 1 | 2 | 4 | +.18 | 2 | 1 | 1 | 4 | 4.19 | ||||||||||||||
| Totals or means | 2 | 1 | 1 | 2 | 7 | 3 | 5 | 4 | 2 | 3 | 2 | 2 | 1 | 35 | +.49 | .77 | 6 | 11 | 6 | 6 | 2 | 31 | 4.43 | .31 |
| Generation. | Grade of offspring. | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lower group. | Upper group. | |||||||||||||
| 3¼ | 3½ | 3¾ | 4 | Totals. | Means. | Standard deviation. | 5 | 5¼ | 5½ | 5¾ | Totals. | Means. | Standard deviation. | |
| F₁ | 3 | 3 | 6 | 3.87 | 6 | 4 | 10 | 5.60 | ||||||
| F₂ | 1 | 1 | 2 | 3.37 | 2 | 3 | 15 | 10 | 30 | 5.52 | ||||
| F₃ | 1 | 7 | 3 | 11 | 5.55 | |||||||||
| Totals or means | 1 | 1 | 3 | 3 | 8 | 3.75 | .25 | 2 | 4 | 28 | 17 | 51 | 5.54 | .19 |
| Grade of (F₁) parents. | Grade of (F₂) offspring. | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lower group. | Upper group. | |||||||||||||||||
| 3 | 3¼ | 3½ | 3¾ | 4 | 4¼ | 4½ | Totals. | Means. | Standard deviation. | 5 | 5¼ | 5½ | 5¾ | Totals. | Means. | Standard deviation. | ||
| Lower group parents | 3⅝ | 2 | 1 | 10 | 8 | 1 | 22 | 3.56 | ||||||||||
| 3¾ | 1 | 2 | 2 | 7 | 2 | 1 | 15 | 3.92 | ||||||||||
| 3⅞ | 1 | 2 | 6 | 8 | 4 | 1 | 22 | 3.92 | ||||||||||
| Totals or means | 2 | 3 | 14 | 16 | 16 | 6 | 2 | 59 | 3.78 | .33 | ||||||||
| Upper group parents | 5¼ | 1 | 1 | 3.50 | 2 | 2 | 5.50 | |||||||||||
| 5⅜ | 1 | 1 | 3 | 1 | 4 | 10 | 3.90 | 2 | 6 | 49 | 13 | 70 | 5.51 | |||||
| 5⅝ | 2 | 5 | 7 | 5.43 | ||||||||||||||
| Totals or means | 1 | 2 | 3 | 1 | 4 | 11 | 3.86 | .35 | 2 | 8 | 56 | 13 | 79 | 5.50 | .15 | |||
| Grade of F₁ parents. | Grade of (F₂) offspring, lower group parents. | Totals. | Means. | Standard deviation. | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -2 | -1¾ | -1 | ¾ | ½ | ¼ | 0 | +¼ | +½ | +¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 2¼ | 2½ | 3¼ | ||||
| ¾ | 1 | 1 | 1 | 3 | .58 | ||||||||||||||||
| ⅝ | 1 | 1 | 1 | 3 | 1 | 1 | 2 | 1 | 11 | .57 | |||||||||||
| ⅛ | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 9 | .80 | |||||||||||
| 0 | 1 | 3 | 3 | 3 | 1 | 4 | 1 | 1 | 3 | 2 | 22 | .72 | |||||||||
| +¼ | 1 | 1 | 1 | 3 | 1.00 | ||||||||||||||||
| +½ | 1 | 1 | 1 | 1 | 4 | -.12 | |||||||||||||||
| +1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 9 | .19 | |||||||||||
| Totals or means | 2 | 1 | 4 | 1 | 6 | 4 | 9 | 1 | 2 | 2 | 9 | 4 | 4 | 2 | 5 | 1 | 3 | 1 | 61 | .58 | 1.17 |
| Grade of F₁ parents. | Grade of (F₂) offspring, upper group parents. | Totals. | Means. | Standard deviation. | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -2 | -1¾ | -1 | ¾ | ½ | ¼ | 0 | +¼ | ½ | ¾ | 1 | 1¼ | 1½ | 1¾ | 2 | 3½ | 4 | 4¼ | 4½ | 4¾ | 5 | 5¼ | 5½ | L[1] | U[2] | L[1] | U[2] | L[1] | U[2] | |
| 4⅛ | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 3 | 5 | 4.85 | |||||||||||||||||||
| 4¼ | 1 | 1 | 6 | 1 | 1 | 1 | 1 | 10 | 4.62 | ||||||||||||||||||||
| 4⅜ | 1 | 1 | 2 | 1 | 1 | 2 | 4 | 4.56 | |||||||||||||||||||||
| 4½ | 1 | 2 | 2 | 1 | 1 | 2 | 4 | 10 | 6 | 8 | 4 | 6 | 6 | 41 | 4.78 | ||||||||||||||
| 4⅝ | 1 | 1 | 1 | 1 | 1 | 2 | 3 | 2 | 8 | 4.91 | |||||||||||||||||||
| Totals or means | 1 | 1 | 2 | 4 | 2 | 1 | 3 | 1 | 2 | 9 | 18 | 10 | 12 | 9 | 7 | 14 | 68 | +.75 | 4.77 | 1.03 | .44 | ||||||||
Top row, a set of arbitrary grades used in the classification of the rats studied.
Middle and bottom rows, skins of rats graded as indicated by the numerals above each skin. The animals graded, +4, +4¼ and +4¾, being entirely dark above, are shown in ventral view.
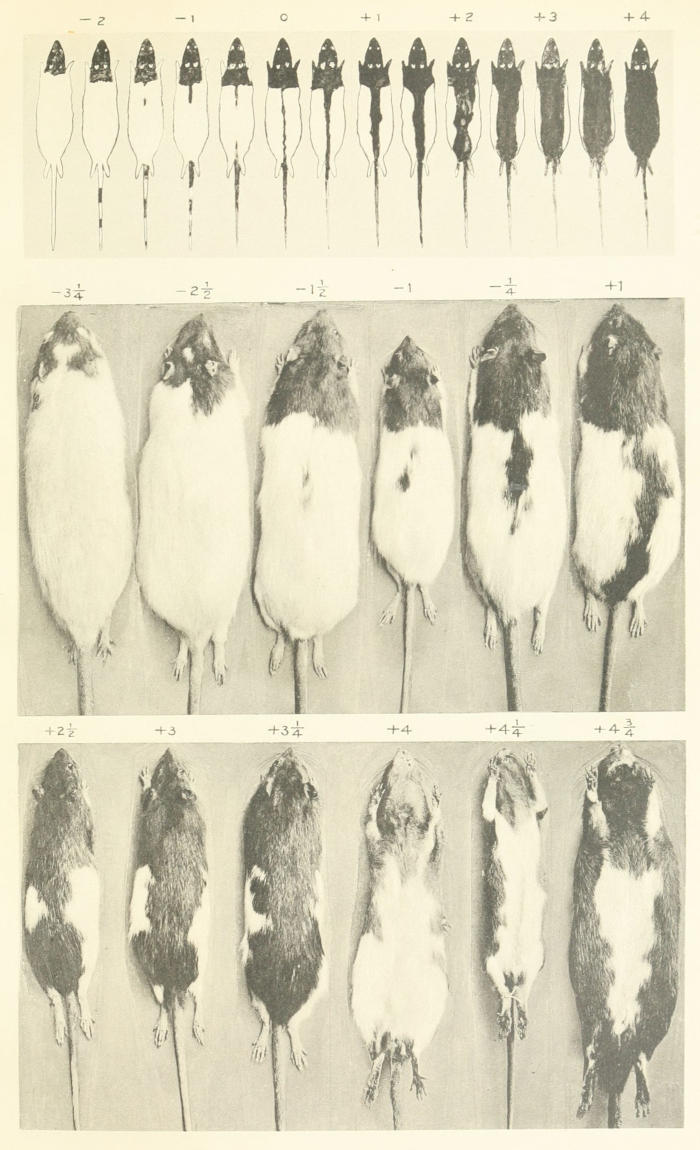
PLATE 1
♂ 8000, a wild gray rat caught in Cambridge, Mass., October, 1908.
♀ 6176, a black hooded rat of grade -1¾ which was mated with ♂ 8000.
♂ 8021 and ♂ 8018, F₁ offspring of the pair just described, ♂ 8000 and ♀ 6176. ♂ 8021 is of grade +4¾, with considerable white below. Notice also his white legs and compare with those of his father. ♂ 8018 is of grade +5½. Notice white areas on belly and front legs.
8075-8078, four F₂ progeny (grandchildren) of ♂ 8000 and ♀ 6176. 8075 is black hooded, grade ¾; 8076 is gray, +4¼; 8077 is gray, +4; 8078 is black, +5½.
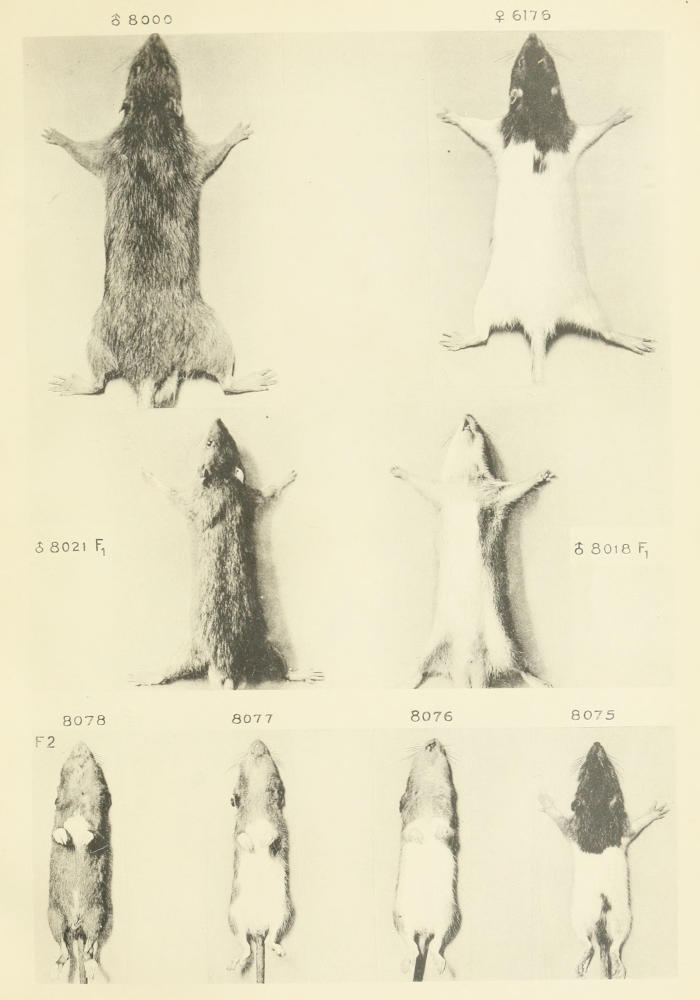
PLATE 2
♂ 103, an evenly marked rat of grade +3. A female rat of this same grade was mated with ♂ 8000 (the wild gray male). Two litters of F₂ grandchildren are shown in 8062-8067, and 8070-8074.
8062 is black hooded, +2; 8064 is gray hooded, +2; 8065 is black, +5; 8066 is gray, +5¼; 8067 is gray, +4½ (notice white feet).
8070 is black hooded, +2; 8071 is gray hooded, +1¾; 8072 is gray, +5; 8073 is black, +5½; 8074 is gray, +5¾.

PLATE 3