
Title: The Beaked Whales of the Family Ziphiidae
Author: Frederick W. True
Release date: January 29, 2014 [eBook #44785]
Language: English
Credits: Produced by Chris Curnow, Joseph Cooper, Stephen Hutcheson,
The Internet Archive/American Libraries and the Online
Distributed Proofreading Team at http://www.pgdp.net
SMITHSONIAN INSTITUTION
UNITED STATES NATIONAL MUSEUM
Bulletin 73
BY
FREDERICK W. TRUE
Head Curator, Department of Biology, U. S. National Museum
WASHINGTON
GOVERNMENT PRINTING OFFICE
1910
BULLETIN OF THE UNITED STATES NATIONAL MUSEUM
Issued September 28, 1910.
The scientific publications of the United States National Museum consist of two series, the Proceedings and the Bulletins.
The Proceedings, the first volume of which was issued in 1878, are intended primarily as a medium for the publication of original, and usually brief, papers based on the collections of the National Museum, presenting newly-acquired facts in zoology, geology, and anthropology, including descriptions of new forms of animals, and revisions of limited groups. One or two volumes are issued annually and distributed to libraries and scientific organizations. A limited number of copies of each paper, in pamphlet form, is distributed to specialists and others interested in the different subjects as soon as printed. The date of publication is printed on each paper, and these dates are also recorded in the tables of contents of the volume.
The Bulletins, the first of which was issued in 1875, consist of a series of separate publications comprising chiefly monographs of large zoological groups and other general systematic treatises (occasionally in several volumes), faunal works, reports of expeditions, and catalogues of type-specimens, special collections, etc. The majority of the volumes are octavos, but a quarto size has been adopted in a few instances in which large plates were regarded as indispensable.
Since 1902 a series of octavo volumes containing papers relating to the botanical collections of the Museum, and known as the Contributions from the National Herbarium, has been published as bulletins.
The present work forms No. 73 of the Bulletin series.
Richard Rathbun,
Assistant Secretary, Smithsonian Institution,
In charge of the United States National Museum.
Washington, D. C., June 1, 1910.
By Frederick W. True,
Head Curator, Department of Biology, U. S. National Museum.
The beaked whales belonging to the family Ziphiidæ are, with the exception of the bottle-nosed whales of the genus Hyperoödon, among the rarest of cetaceans. Of the three genera Mesoplodon, Ziphius, and Berardius, so far as I have been able to ascertain from published records, specimens representing about one hundred individuals are known, and somewhat more than one-half of these belong to the first-named genus. Berardius is the rarest genus, only about fourteen specimens having been collected thus far. The U. S. National Museum contains specimens representing some twenty-five individuals of the three genera, or about one-fourth of the material at present available. Among these are six specimens of the genus Berardius, or nearly half of all that have been recorded thus far.
The most important addition to the knowledge of these whales made during the last quarter century was the discovery of representatives of the three genera Mesoplodon, Ziphius, and Berardius, at Bering Island, in the North Pacific, by Dr. Leonhard Stejneger, whereby the known range of the family was very greatly extended. Two of the forms were described by Doctor Stejneger in 1883, and the third by myself from a skull which he collected. About one-half of the material which the Museum possesses consists of that collected by Doctor Stejneger in Bering Island and that from the same locality presented by Mr. Nicholas Grebnitzki, Russian governor of the Commander Islands.
About six years ago the National Museum received information and specimens from correspondents showing that the range of the three genera found at Bering Island extends to the eastern North Pacific, one genus (Ziphius) having been observed at Kiska Harbor, Alaska, another (Mesoplodon) at Yaquina Bay, Oregon, and the third (Berardius) at St. George Island, Pribilof Group, Alaska, and near Cape Mendocino, California.
On the east and west coasts of the United States the only occurrences of beaked whales known to me are as follows:
Of this genus the National Museum has four specimens; namely, (1) a skull (Cat. No. 21112, U.S.N.M.) obtained at Bering Island, North Pacific Ocean, in 1883, by Dr. L. Stejneger, and made the type of the species M. stejnegeri True; (2) a skull and photographs (Cat. No. 143132, U.S.N.M.) of the same species, from Yaquina Bay, Oregon, obtained in exchange from Mr. J. G. Crawford in 1904; (3) a skeleton, cast, and photographs of a young male (Cat. No. 23346, U.S.N.M.), hitherto supposed to represent M. bidens, caught at Atlantic City, New Jersey, in 1889; and (4) a skeleton of an adult (Cat. No. 49880, U.S.N.M.) from the Chatham Islands, New Zealand, representing M. grayi.[1]
In addition to this material, I have had the privilege of examining two skulls belonging to the Museum of Comparative Zoölogy, and hitherto supposed to represent M. bidens, and two skeletons belonging to the American Museum of Natural History. Of these last, one is that of an adult and was purchased by the American Museum under the name of M. layardi, but was subsequently recognized to be a new species and was described by Mr. Andrews, under the name of Mesoplodon bowdoini. The other is that of a young individual, and has been labeled M. grayi.
As already noted by Dr. G. M. Allen,[2] only four specimens of Mesoplodon have been recorded hitherto from the Atlantic coast of the United States. These are:
1. An adult, sex unknown, but probably female, 16 feet long, found at Nantucket, Massachusetts, in 1867, and recorded by Prof. L. Agassiz.[3] The skull of this individual is in the Museum of Comparative Zoölogy, Cambridge, Massachusetts.
2. A young male, 12½ feet long, captured at Atlantic City, New Jersey, March 28, 1889. The skeleton (Cat. No. 23346, U.S.N.M.) is in the National Museum.
3. A young female, 12 feet 2 inches long, stranded at Annisquam, Massachusetts, August, 1898, and recorded by the late Alpheus Hyatt.[4] The skeleton is in the museum of the Boston Society of Natural History.
4. An adult female, said by fishermen who measured it to have been 22 feet long, entangled in pound nets at North Long Branch, New Jersey, July 22, 1905, and recorded by Dr. Glover M. Allen.[5] The cranium of this individual is preserved in the Museum of Comparative Zoölogy. The rostrum and mandible, which were originally obtained, were afterwards destroyed by accident.
I have examined all this material. Writers who have had occasion to mention these four specimens thus far have referred them tacitly to Mesoplodon bidens (Sowerby), but, after a careful study of them, I have ascertained that while the Nantucket specimen belongs to that species, the Atlantic City and Long Branch [4] specimens represent Mesoplodon europæus (Gervais). This is a very interesting discovery, because the latter species has been known hitherto only from a single skull, and its validity has been frequently questioned. The Annisquam specimen, as will be seen later, presents characters which appear to ally it to M. densirostris.
The only specimen from the Atlantic coast of the United States which can with certainty be referred to this species is the one from Nantucket mentioned on page 3. Prof. L. Agassiz’s original notice of it is so brief that it is quoted in full below:
Professor Agassiz also brought to the notice of the Society the discovery of a Cetacean, new to America. The skull was exhibited, and its peculiar features pointed out. It was obtained on the coast of Nantucket by Messrs. H. M. and S. C. Martin, of Roxbury. It belonged to the genus Mesoplodon, as characterized by Gervais, and ought to be separated from the fossil Ziphius, described by Cuvier. Professor Agassiz, however, questioned whether Mesoplodon was not identical with Delphinorhynchus, previously described by De Blainville. The specimen found at Nantucket measured 16 feet in length.[6]
The skull of this Nantucket specimen, which I have before me, is thoroughly adult. That the specimen is a female is probable from the fact that the teeth (one of which is preserved), though fully developed, are only two-thirds as broad and three-fourths as long as those of Sowerby’s specimen (the type of the species), which was an adult male.[7] The skull is 765 mm. long, and about 30 mm. are lacking from the end of the beak, so that the original length was about 795 mm. It appears to be, therefore, rather the largest skull of the species of which there is any record. The specimen itself, according to Dr. J. A. Allen, was 16 feet 3 inches long.[8] The largest European skull appears to be the one in the Edinburgh Museum, described by Sir William Turner in 1872.[9] The length of this is 749 mm. The specimen was a female, but though the skull is so large, the mesirostral cartilage was not ossified, and the individual was, therefore, probably not thoroughly adult. Two other European specimens, of which the total length was almost identical with that of the Nantucket specimen, were (1) the adult female obtained at Overstrand, England, in 1892, and recorded by Southwell and Harmer[10] (length 16 feet [5] 2 inches, straight); (2) the adult male obtained at Brodie House, Scotland, in 1800, and recorded by Sowerby[11] (length 16 feet). The length of the skull is not given for either of these specimens. The adult male obtained at Rugsund, Norway, in 1901, and recorded by Grieg,[12] was only 15 feet 1 inch long, but some of the measurements of the skull are as large as, or even a little larger than, those of the Nantucket skull. The total length of the skull was not given, as the end of the beak was lacking.
Grieg’s figures of the Rugsund skull afford a very satisfactory basis for comparisons between that specimen and the Nantucket skull (Pl. 1, fig. 1). Both skulls show the comparatively narrow frontal region, the moderately developed tubercle anterior to the anteorbital notch, and the low maxillary ridge, which are characteristic of the species. In both skulls the anterior prolongation of the ethmoid is lanceolate and flat, but in the Rugsund skull the apex is truncated. In the latter also the posterior end of the mesirostral ossification is divided into three longitudinal sections by two lateral and somewhat divergent grooves, while in the Nantucket skull there is only a single median groove. These differences may safely be regarded as individual. Toward the distal end the surface of the ossification in the Nantucket is pitted and irregular and descends much below the level of the premaxillæ. It ends distally at the same point with the vomer. In this skull the proximal end of the premaxillæ and adjoining plate of the maxillæ are somewhat less reflexed than in the Rugsund skull. The shape of the superior margin of the supraoccipital is alike in both.
There are no well-defined differences in the relative thickness of the beak at the base or in the form and position of the visible portion of the palatines, but in the Nantucket skull the mass of the combined frontal and lachrymal anterior to the orbit is less rounded and more triangular than in the Rugsund skull. The temporal fossæ also have a postero-superior angular enlargement not seen in the latter.
In the Nantucket skull the rostral portion of the premaxillæ is high and at the distal end vertical. The superior profile is somewhat convex, and the superior free margin rounded proximally, but sharp distally. The least distance between the free margins is 10 mm.
The pterygoids are cut off from the maxillæ anteriorly by a very narrow band of the palatine, which connects with a broad band externally and a lanceolate segment internally. The inferior pterygoid ridges diverge anteriorly. The broad surface internal to them is concave. The external border of the pterygoid sinus is nearly straight. An elongated, fusiform section of the vomer is visible on the inferior surface of the beak at the middle for a distance of 158 mm., and a small lozenge-shaped section, ill defined, is visible between the pterygoids and palatines. (Pl. 4, fig. 1.)
The expanded anterior end of the malar is rhomboidal in form, with an external free margin 11 mm. long. Anteriorly it does not form part of the margin of the anteorbital notch.
The lachrymal is irregularly oblong, with an external free margin 35 mm. long and 12 mm. thick. The distance from the anteorbital notch to the anterior end of the orbit is 60 mm. (Pl. 7, fig. 1.)
The lateral free margins of the basioccipital are extended posteriorly beyond the exoccipitals, which is a character indicative of age.
The supraoccipital has a distinct median ridge, with a longitudinal depression on each side, bounded externally by a prominent convexity. (Pl. 10, fig. 1.)
The mandible is slender, with a very elongate symphysis, which measures 237 mm. The inferior outline of the ramus is strongly concave at the middle and slightly convex posteriorly, while the symphysial portion is bent upward. The superior outline is concave both behind and before the tooth, and also immediately anterior to the coronoid process. At about the beginning of the posterior fourth the outline is convex, and the mandible at this point is nearly as deep as at the coronoid process. The superior surface of the symphysis slopes down on each side to the median line, but each half of the surface is itself nearly plane. (Pl. 11, figs. 1, 2, and 5.)
The alveolar groove anterior to the tooth is very distinct throughout and is without septa and open at the bottom. It ends distally in a rounded aperture 6 mm. in diameter, below which are several small foramina. These lead to a very large canal which occupies all the symphysial portion of the mandible, the walls being comparatively thin. Behind the tooth the alveolar groove becomes narrower gradually and disappears in a length of about 140 mm.
The mental foramen is situated in line with the anterior base of the tooth, and is confluent with a groove which extends forward for about 80 mm. A rather shallow groove runs along the inferior margin of the symphysis.
The coronoid process is erect and rounded, and is joined by a horizontal ridge anteriorly.
The mandibular tooth, which is shown in Pl. 2, fig. 3, is preserved on the right side only. Its dimensions are as follows: Length anteriorly in a straight line, 75 mm.; length from the apex to the posterior end of the root, straight, 60; greatest antero-posterior breadth, 28; transverse thickness, 10; height of apex above internal superior margin of jaw when tooth is in situ,[13] 22; antero-posterior length of base of exposed portion, 30; distance from anterior end to posterior end of root, 37; greatest height of the exposed dentine crown, above the cement, 14; length of the base of the dentine crown, 12.
This tooth, as already stated, is only two-thirds as broad and three-fourths as long as that of Sowerby’s Brodie House specimen (the type of the species), which was an adult male, and leads to the belief that the Nantucket specimen was a female. This is in a manner confirmed by the Rugsund specimen, which was an adult male and had teeth as large as Sowerby’s specimen. It has to be remarked, [7] however, that in the Overstrand, England, specimen (1892), which was an adult female, the teeth did not project beyond the gums. Messrs. Southwell and Harmer say regarding it:
The jaws were apparently completely edentulous, and although it was possible to feel through the gums a slight prominence on either side in the position of the teeth of the male, we could not by this means definitely satisfy ourselves with respect to this point, nor were we able to ascertain the presence of any other rudimentary teeth in either jaw. The evidence which exists on this subject is favourable to the view that the female of this species is not provided with any teeth which are large enough to pierce the gums.[14]
It is probable that the teeth in the Nantucket specimen, though quite large, did not project beyond the gums any considerable distance. The external border of the alveolar groove behind the tooth is only 20 mm. below the apex of the tooth, and it is not unlikely that the gums in a specimen of this size had nearly that thickness, so that only the tip of the tooth would project beyond them. Though the apex is acute, it has a flat abraded surface anteriorly, which, however, is but 4 mm. long. It seems probable, on the whole, that the teeth in the female may be quite large without projecting more than a few millimeters beyond the gums.
In shape the tooth of the Nantucket specimen is almost identical with that of Sowerby’s Brodie House adult male, as figured by Lankester. The dentine at the apex is more nearly white than the cement which surrounds it. The superior margin of the latter is not a plain ring, but sends upward a papilliform projection on each side. The dentine itself has two vertical grooves on each side. The root of the tooth ends very obliquely and is rugose and irregular. The cavity is closed.
Grieg remarks as follows regarding the structure of the teeth of the Rugsund specimen:
Sections and microscopic preparations of the alveolar tooth of this whale show that its apex consists of dentine, within which is found an inner pulp cavity 4 mm. long and 1 mm. broad. The dentine, the structure of which agrees with that which Turner found in Mesoplodon bidens and Mesoplodon layardi, is yellowish white, with the exception of the part nearest the pulp cavity, which is yellowish brown. It seems to correspond most closely to what Ray Lankester called osteodentine. Throughout the tooth the dentine is covered with a very thin layer of shining white enamel. The enamel is, however, lacking on the front of the tooth, having probably been worn away. A section through the middle of the tooth, at right angles with the V-shaped furrow, shows a yellowish cement layer from 3 to 5 mm. broad, which is, however, worn away on the front of the tooth. Within the cement layer is a white, amorphous, calcareous mass, forming a band from 1.5 to 3.5 mm. broad, which appears to correspond to Ray Lankester’s “globular matter” and Turner’s “modified vasodentine.” The mass seems to agree most closely with Ray Lankester’s “globular matter,” as it has “no structure excepting an indistinct botryoidal character visible with a low magnifying power.” The core of the tooth consists of dentine, the inner layer of which is brownish, while the outer is rather whitish yellow. As above mentioned, the dentine is visible on the front of the tooth, since both the cement and the amorphous, calcareous mass are worn away. Moreover, it is clear that on the front of the tooth the dentine is not covered by enamel. The pulp cavity is reduced to a fine pore. A section across the root of the tooth shows an outer yellowish cement layer, from 2 to 5 mm. broad, while the interior of the tooth is filled with a white, amorphous, calcareous mass, which is interspersed with thin yellowish lamellæ of dentine. Here and there, also, thin lamellæ are seen to extend from the outer cement layer into the white, amorphous, calcareous mass. The dentine lamellæ appear to be identical with what Ray Lankester calls osteodentine. No pulp cavity is visible in the root of the tooth.[15]
The dimensions of the Nantucket skull are given in the following table in comparison with those of seven European skulls of M. bidens. Dimensions of the Annisquam, Massachusetts, skull are also added for purposes of comparison, although it represents another species (see p. 9).
Dimensions of eight skulls of Mesoplodon bidens and one skull of M. densirostris (?).
| Measurements. | B | C | D | E | F | G | H | I | J |
|---|---|---|---|---|---|---|---|---|---|
| mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | |
| Total length | b765+ | 749 | 620 | 743± | ... | 733 | 740 | 660 | c622 |
| Length of rostrum | b483+ | 489 | 400 | ... | ... | 485 | 500 | 410 | c377 |
| Tip of beak to end of pterygoid | bd607+ | 572 | ... | ... | ... | 582 | 590 | 517 | cd466 |
| Height from vertex to pterygoid | 277 | 241 | ... | 254 | 267 | 272 | 258 | 235 | 248 |
| Breadth between orbits | e277 | 286 | f254 | 267 | 292 | 293 | 253 | f260 | [278] |
| Breadth between zygomatic processes | 289 | 292 | 262 | 292 | 295 | 298 | 270 | 268 | 266 |
| Breadth at maxillary notches | 184 | 197 | 170 | 184 | 193 | 187 | 170 | 175 | [166] |
| Breadth of beak at middle | 42 | 51 | 38 | ... | ... | 36 | 46 | g40 | 38 |
| Depth of beak at middle | 35 | ... | h31 | ... | ... | ... | ... | h33 | 51 |
| Greatest breadth of premaxillæ proximally | 131 | 127 | 115 | 114 | 116 | 129 | 124 | 122 | ... |
| Greatest breadth of premaxillæ in front of anterior nares | 107 | 102 | h104 | 102 | 108 | 108 | 100 | h76 | 92 |
| Greatest breadth of anterior nares | 54 | ... | 53 | ... | 53 | 50 | 50 | 50 | 39 |
| Length of temporal fossæ | 90 | ... | ... | ... | ... | ... | h66 | 82 | |
| Breadth between temporal fossæ | 222 | ... | ... | ... | ... | ... | ... | ... | 208 |
| Breadth of foramen magnum | 50 | ... | ... | ... | 49 | 56 | 54 | 80 | 46 |
| Length of mandible | c651 | ij470 | 543 | i464 | ... | 639 | 640 | 560 | ... |
| Length of symphysis | 237 | 241 | 162 | ... | ... | 212 | 220 | 160 | ... |
| Greatest depth of mandible | 106 | 114 | 92 | 102 | 116 | 110 | 97 | 95 | ... |
The skull of the specimen from Annisquam, Mass., (Pl. 1, fig. 2) is, I regret to say, in rather poor condition. It is broken in the left orbital region, and all the bones, especially those of the beak, are warped by weathering. The proximal extremity of the left premaxilla is lacking and also the tip of the beak.
The skull is obviously that of a young animal, as all the sutures are open and the surface of the occipital condyles is pitted, owing to imperfect ossification.
Although the dimensions of the skull, with a few exceptions, agree well with those of young specimens of M. bidens, as shown by the foregoing table (p. 8), certain differences stand out conspicuously. The most salient of these is the depth of the beak as a whole and the depth and shape of the rostral portion of the premaxillæ. The latter portion of the premaxillæ instead of being low, with a straight inferior margin, is very high, with the inferior margin strongly convex. At the middle of the beak the premaxillæ are higher than the maxillæ on which they rest. It is true that the shape of the beak varies greatly with age in bidens and other species of Mesoplodon, but I do not find any evidence that such a change as is here indicated takes place in bidens. The form of the beak and of the rostral portion of the premaxillæ is that of M. densirostris.
The beak is almost as broad at the base as in bidens, but the lateral free margin of the maxilla anterior to the anteorbital notch instead of continuing along the side of the beak nearly to the tip, as in bidens, ends at a point about 90 mm. in front of the line of the notch, beyond which the sides of the beak are vertical.
The margin of the maxilla immediately anterior to the anteorbital notch is a little damaged, but there was apparently no strong tubercle at this point, and the surface of the maxilla, though convex, is not raised into a distinct ridge. In a young skull, however, one would not expect to find a high ridge. The palatines are visible from above, which is not the case in bidens.
The maxillary foramen is situated a little in advance of the premaxillary foramen and is directed forward, and, as Dr. Glover M. Allen has pointed out, connects with a broad groove which runs forward along the triangular, horizontal portion of the maxilla at the base of the beak. The maxillæ are much broader behind the notch than in bidens, and the anterior end of the malar forms the bottom of the notch. The premaxillæ are noticeably constricted immediately in front of the premaxillary foramina, and the expanded portion just behind these foramina is nearly horizontal, with a low transverse ridge near the middle. The proximal end of the premaxillæ is nearly vertical. The anterior nares are noticeably small. The foramen magnum is large, with a trifoliate outline (Pl. 10, fig. 2). The palate at the proximal end presents a median ridge with a narrow groove on each side. The palatines extend as a broad band much beyond the pterygoids anteriorly. The vomer is visible below for a space of 142 mm. near the end of the beak. A very small piece is also visible at the base of the beak, between the palatines and [10] pterygoids. The inferior surface of the pterygoids is convex on the side adjoining the lateral free margin (Pl. 4, fig. 2).
This skull is peculiar in that there is no very distinct basirostral groove and that the basirostral ridge, as already stated, extends forward only about 90 mm. Below this ridge is a shallow broad groove which narrows rapidly forward and can be traced to the extremity of the beak, where it broadens out somewhat (Pl. 7, fig. 2).
While this skull agrees in size and in many of its proportions with similar skulls of M. bidens, it differs from that species and agrees with M. densirostris in the breadth across the anteorbital region, in the depth of the beak and its shape at the base, in the shape of the premaxillæ both distally and proximally, in the direction of the maxillary foramen, and the shape of the maxillary bone in front of the same, in the occupation of the base of the maxillary notch by the anterior end of the malar, in the absence of any distinct maxillary ridge above the notch, in the forward extension of the palatines, and in the shape of the foramen magnum.
Flower states that there is a deep basirostral groove in M. densirostris,[16] but neither the figure in Gervais’ Zoologie et Paleontologie Française,[17] nor that in Van Beneden and Gervais’ Ostéographie des Cétacés,[18] shows such a groove. The conformation of the base of the rostrum appears to be about the same as in the Annisquam skull.
In regard to differences between this skull and those of M. densirostris it should be stated that in the latter the premaxillary foramina are situated farther apart, and that the maxillary foramina are situated considerably in advance of those of the premaxillæ instead of nearly in line with them.
The Annisquam skull approaches M. europæus in several characters, but these are such as europæus shares with densirostris. The principal ones are the breadth of the maxillæ in front of the orbits, the presence of the malar in the base of the anteorbital notch, and the convexity of a part of the inferior surface of the pterygoids.
Dr. Glover M. Allen has given an account of the exterior, skeleton, and teeth of this specimen, from which the following particulars are extracted:[19]
Regarding the Annisquam specimen no color notes were taken, but from a few small photographs in the possession of the Boston Society of Natural History, it appears evident that the ventral portion was of a lighter tint, and in one of the views a few oval whitish spots are seen on the side a trifle behind the middle portion of the body. Another view shows the convexity of the posterior margin of the flukes at the median point, as well as the prominent dorsal fin. The lower jaw protruded slightly beyond the upper. Measurements of this specimen, as noted by Professor Hyatt, are as follows: Total length, 12 feet 2 inches; from anus to bight of flukes, 3 feet 4 to 6 inches; across flukes, 3 feet 1 inch; from tip of rostrum to angle of mouth, 1 foot 1½ inches. The gular furrows were noted as about 10 inches long and from ¼ to ½ an inch deep.
The teeth of the Annisquam specimen barely projected above the alveoli of the jaws and are sharply mucronate. The basal portion of each, however, is more like that of the male’s tooth [M. europæus] in the slightly convex posterior outline and the forward extension of the anterior angle. * * *
The Annisquam skeleton has 45 vertebræ. Four of the seven cervicals are fused. The atlas, axis, and third cervical are firmly anchylosed throughout, save for the lateral foramina for the passage of the [11] cervical nerves. The fourth cervical is fused to the third by the dorsal spine on the left side and by the tip of the upper lateral process of the same side. Its centrum, right half of the dorsal spine (the spine is divided medially), and the remaining lateral processes are free. * * * The epiphyses of the fourth and fifth cervical vertebræ and the anterior epiphysis of the sixth cervical are fused to their respective centra, but all the other epiphyses of the vertebral column and of the pectoral limbs are free.
The Annisquam skeleton has nine dorsal vertebræ with their corresponding pairs of ribs. * * * The sternum of this specimen presents few points of interest. It consists of four pieces, the anterior-most of which is largest, slightly hollowed above, and correspondingly convex below. The three remaining pieces are nearly flat, with a deep median notch at the anterior and posterior border of each. The posterior piece evidently represents a fusion of the elements of two segments, as there are articular surfaces for two pairs of ribs.
From the foregoing, it appears that the Annisquam specimen probably had one or two vertebræ less than bidens or europæus, and that the sternum was somewhat differently shaped. The tooth, which is figured by Doctor Allen, is conical, compressed, 54 mm. long, 30 broad at the base, and resembles teeth of immature bidens.
Although with such scant material it is not possible to determine satisfactorily the identity of this third species of Mesoplodon in the North Atlantic, represented by the Annisquam specimen, I feel convinced that that specimen does not belong to M. bidens and that there is a strong probability that it belongs to M. densirostris. It is true that the latter species has been found hitherto only in the Indian Ocean and about Australia, but we know so little about the distribution of the ziphioid whales that, in my opinion, that circumstance by itself should not be given very great weight.
This species was based on a single specimen found floating in the English Channel about seventy years ago. An account of the circumstances under which it was found was given by Eugène Deslongchamps in 1866, as follows:
The head, which forms the subject of this last note, was given to my father some twenty-five or thirty years ago by Mr. Abel Vautier, a merchant and armorer of our town, who died at Paris two years since.
The captain of one of Mr. Vautier’s ships, on his return from a voyage to the colonies, saw floating on the water, at the entrance to the English Channel, the body of a large animal entirely covered by birds (large and small gulls, etc.), which were devouring it. The ship approached the stray, and the captain, knowing that Mr. Abel Vautier was greatly interested in natural objects, had the head of the cetacean cut off, fastened it securely with a cord, and let it trail behind the ship. When he arrived at Caën he made a present of it to Mr. Vautier. The piece had at that time an appearance anything but agreeable. Mr. Vautier was especially fond of beautiful objects which please the eye, and hence he offered it to my father, saying, “You, who are an anatomist, can make better use of this than I can.” My father was unwilling to refuse the present, but neither he nor Mr. Vautier knew as yet of its extreme rarity. It is in fact, up to the present time, the only specimen which exists, and is a unique object in collections.[20]
No additional specimens have been recorded from European waters or elsewhere, and much doubt has been thrown on the validity of the species, many zoologists regarding it as an adult of the commoner species M. bidens. Van Beneden remarked in 1888:
The opinions of naturalists are divided as regards the identity of this ziphioid, which is unique up to the present time. In the eyes of some it represents an old male of the common Mesoplodon, in which the tooth, instead of developing near the middle of the jaw, has developed near the anterior extremity. This is the opinion of Doctor Fischer and others, who think that this unique specimen represents merely an individual modification and that consequently it should not figure in the list of species. We do not share this opinion. It is not impossible that this ziphioid may belong to the other hemisphere, and this would explain why only one single individual has been captured in Europe.[21]
In view of the circumstances surrounding the discovery of the original specimen, it is of great interest to find that two of the specimens from the east coast of the United States represent the same species. As one of them is adult and the other young, the view that the type of M. europæus is merely an old individual of M. bidens is satisfactorily disposed of, as is also the opinion that it represents a singular individual variation.
The two American specimens which represent europæus are those from North Long Branch, New Jersey (adult female; skull, lacking rostrum and mandible, in the Museum of Comparative Zoology), and from Atlantic City, New Jersey (young male; skeleton, cast and photographs in the U. S. National Museum, Cat. No. 23346).
The species europæus differs from bidens in the following characters, which may be regarded as diagnostic:
Size larger and pectoral limbs relatively shorter and narrower.
The expanded portion of the maxillæ and frontals broader in front of the orbit. The protuberance which projects into the anteorbital notch much larger and the ridge on the maxilla which extends backward from it much higher. Distance from inner margin of maxillary foramen to tip of protuberance much more than one-half the distance between the maxillary foramina of the two sides. Rostrum deeper at the base. Inferior surface of pterygoids more or less convex, with a ridge (in adults) running diagonally across it.
The cranial characters above enumerated are found in the type-skull, as will be seen by examining the excellent figures in Van Beneden and Gervais’ Osteography, plate 24.
In Dr. Glover M. Allen’s account of the Long Branch specimen[22] it is stated that the fishermen who measured it reported that it was 22 feet long, while none of the European specimens (some of which were certainly adults) was more than 16½ feet long. That the measurement reported by the fishermen is at least approximately correct appears from the fact that the skull is larger than that of any of the European specimens. The beak is missing, so that the total length of the skull can not be given, but the distance from the occipital condyles to the line of the maxillary [13] notches (straight) is 312 mm., while in the largest adult among the European specimens this distance is only 260 mm., and in the thoroughly adult Nantucket specimen 282 mm.
The Atlantic City and Long Branch skulls also agree in numerous other details of structure in addition to the foregoing, the more important of which will now be mentioned. Unless otherwise stated, the type-skull, as shown by Van Beneden and Gervais’ figures,[23] also presents the same peculiarities in contrast with M. bidens.
Dorsal aspect (Pl. 2, figs. 1 and 2).—The premaxillæ are more depressed immediately in front of the blowhole than in M. bidens, which, with the prominence of the maxillary ridges, makes this whole region appear strongly concave. The blowhole is narrower absolutely and also relatively to the breadth of the expanded proximal ends of the premaxillæ, so that while in bidens the breadth of the blowhole is much more than one-third the breadth across the proximal ends of the premaxillæ, in europæus it is considerably less than a third. Both premaxillæ are much constricted on the sides of the blowhole and the effect is heightened by the greater expansion of the proximal ends of the former. These ends do not fit closely against the adjoining edge of the maxillæ as in bidens, but leave a transverse vacuity, or trough, which is especially noticeable in the type-skull. The anterior end of the malar bone occupies the bottom of the maxillary notch and a small portion of it is visible from above, while in bidens it does not extend up into the notch at all from the inferior surface and is not visible from above. The posterior margin of the maxillæ is more squared in europæus than in bidens.
The margins of the beak, formed by the maxillæ, instead of being straight, are somewhat emarginate a little posterior to the middle of the length and somewhat convex anterior to it, which gives the contour of the beak, seen from above, a different shape from that of bidens. In the type-skull of europæus the mesirostral ossification appears to be higher at the proximal end than the premaxillæ, and distally extends to the end of the beak. In bidens it is lower than the premaxillæ and, in the Nantucket skull at least, ends anteriorly at the same point as the vomer, or, in other words, much behind the end of the beak. It would appear from the statements of Sir William Turner, Van Beneden and Gervais, Grieg, and others, that the mesirostral ossification never reaches the end of the beak in bidens, but it does in grayi, haasti, densirostris, and many fossil species, as well as in europæus.
Lateral aspect (Pl. 8, figs. 1, 2).—The temporal fossæ are a little longer than the orbit in europæus, but a little shorter than the orbit in bidens; in the former the superior margin is flat or a little concave, rather than convex. The exoccipital extends in an angle farther forward in europæus, and the suture between it and the zygomatic is, in consequence, less nearly vertical than in bidens. The premaxillæ at the sides of the blowhole are nearly horizontal, so that their superior surface is little seen from this aspect, while in bidens they slope downward, so that the whole of the superior surface is visible. The high maxillary ridge, situated behind the anteorbital notch, is very noticeable from this point of view, as it shuts off a [14] considerable portion of the premaxillæ. The convex inferior outline of the beak and its great depth at the base are also salient peculiarities.
Ventral aspect (Pl. 5, figs. 1, 2).—The anterior ends of the palatine bones are bifurcated, the inner part being the smaller. The two bones make but a narrow angle with the median line, instead of a wide one, as in bidens, and the surface of the maxillæ between them is strongly convex instead of flat. This convexity is narrowed at both ends, or, in other words, is fusiform in shape. No similar conformation is found in bidens, in which the inferior basal area of the maxillæ is flat.
In the young Atlantic City skull of europæus, the vomer is visible as a small, narrow, club-shaped piece, 68 mm. long. Anteriorly it joins the premaxillæ, which form a prominent ridge in the median line. On each side of this ridge is a wide and quite deep groove. As the beak is lacking in the adult North Long Branch skull, its peculiarities can not be made known. In the type-skull the form is the same as in the Atlantic City skull, but the vomer does not appear at all on the palate. In bidens the shape of the inferior surface of the premaxillæ at the distal end is quite different. A very narrow groove runs parallel with and close to the median line and the whole surface external to it is more or less convex.
The mandible of the Atlantic City specimen of M. europæus resembles that of the type, as figured by Van Beneden and Gervais, in the shortness of the symphysis and in the position of the tooth, which is in advance of the posterior end of the symphysis. A number of differences, however, require consideration. (Pl. 11, figs. 3 and 6.)
In the type, the symphysis, as shown by Van Beneden and Gervais’ figure, plate 24, fig. 2a, is a little more than one-fifth the length of the mandible. The same relative proportion is found in the Atlantic City specimen, but, as the latter is a younger individual, one would expect the symphysis to be shorter. The figure of Van Beneden and Gervais gives the impression that in the type the end of the mandible is broken, and that, hence, the symphysis is shorter than it was originally. It will be observed that figures 2 and 2a do not agree as regards the length between the tooth and the end of the jaw, figure 2a showing a greater length. In figure 2, however, the jaw seems rather too long for the cranium, and if the greater length of the symphysis shown in figure 2a were introduced, it would certainly be so. The explanation of this discrepancy is not readily found; but one may be allowed to think that the symphysis is not so blunt in the type as is shown in figure 2.
In the Atlantic City specimen the superior lateral free margin of the symphysis is straight, while in the type it is much elevated. This is no doubt due to difference in age and possibly in sex. The type shows three or four mental foramina, while the Atlantic City specimen has one large posterior one and seven smaller ones anterior to it.
Another peculiarity of the latter specimen is that the coronoid process is situated much in advance of the condyle, while the angle extends considerably behind it. In the type both are nearly in line with the condyle. I am unable to explain this difference.
In the Atlantic City specimen the axis of the tooth where it emerges from the alveolus is 91 mm. from the end of the jaw. The portion of the tooth above the alveolus is 11 mm. long at the base and 12 mm. high. It is conical and sharp pointed, and is inclined forward and a little outward, especially at the tip. At the alveolus the transverse breadth of the tooth is 5 mm. The much larger tooth in the type indicates that that specimen was a male.
The mandible of the Atlantic City specimen of M. europæus differs from that of M. bidens in the relative shortness of the symphysis, the large number of mental foramina, the more anterior position of the tooth, and the direction of the crown, which is forward instead of backward.
Dimensions of the type and two other skulls of Mesoplodon europæus.
| Measurements. | A | B | C |
|---|---|---|---|
| mm. | mm. | mm. | |
| Total length | 762 | (b) | 675 |
| Length of rostrum | 459 | ... | 427 |
| Tip of beak to posterior end of pterygoids | 561 | ... | 525 |
| Height from vertex to end of pterygoids | c292? | 283 | 256 |
| Breadth between orbits | 327 | d325 | d287 |
| Breadth between zygomatic processes | 360 | e325 | 302 |
| Breadth at anteorbital notches | 210 | 205 | f182 |
| Breadth of beak at middle | 66 | ... | 60 |
| Depth of beak at middle | 54 | ... | 40 |
| Greatest breadth of premaxillæ proximally | 168 | 147 | 142 |
| The same, in front of anterior nares | 111 | 99 | 104 |
| Breadth of anterior nares | 51 | 45 | 42 |
| Length of temporal fossæ | 102 | 115 | 101 |
| Breadth between temporal fossæ | 228 | 212 | 208 |
| Breadth of foramen magnum | 42 | 34 | 34 |
| Length of mandible | 654 | ... | 565 |
| Length of symphysis | 135 | ... | 116 |
| Greatest depth of mandible | 120 | ... | 101 |
The vertebral formula of three specimens of M. bidens and of the Atlantic City specimen of M. europæus is as follows:
| M. europæus. | ||||
| Atlantic City | C. 7; | Th. 9; | L. 11; | Ca. 20=47 |
| M. bidens. | ||||
| Landenæs | 7; | 10; | 11; | 19=47 |
| Fæø | 7; | 9; | 11; | 19=46 |
| Udsire | 7; | 10; | 9; | 20=46 |
Although the skeleton of M. europæus appears from the foregoing formula to include one less thoracic vertebra than those of M. bidens, as the last pair of ribs present is as long as the preceding ones, an additional pair probably existed originally. The formula for europæus would then be: C. 7, Th. 10, L. 10, Ca. 20 = 47. (Pl. 13, fig. 1.)
In the Atlantic City specimen all the epiphyses are free. The atlas and axis are anchylosed together, the third cervical is united to the axis by the centrum, and on the right side by the top of the neural arch; on the left side the arch is imperfect and free. The fourth to the seventh cervicals, inclusive, are all free. The arch is incomplete above in the fourth, fifth, and sixth, but complete in the seventh. There is a short neural spine on both sixth and seventh cervicals. The atlas has a broad, obliquely-truncated inferior lateral process, but no superior process, while the axis has both inferior and superior processes. The inferior process is twice as long as the superior process, and both are directed backward. They do not meet to form a ring. The third to the sixth cervicals, inclusive, have inferior processes only, that on the third being long and thin (but developed on the left side only). On the fourth and fifth cervicals the processes are short and small; on the sixth, long and broad, and directed downward. The centrum of the seventh cervical has a broad facet on the side, where the first rib is attached, and an inferior lateral process thicker than that of the sixth cervical, but also directed downward.
It is doubtful whether the foregoing characters of the cervical vertebræ are of any systematic importance, as there is a very large amount of individual variation among these animals in the development of the transverse processes and other details of structure. M. bidens, however, appears to have superior transverse processes on most of the cervicals which sometimes unite with the inferior processes to form foramina. In the specimen of M. europæus under consideration there are no superior processes, except on the axis.
Metapophyses are first distinguishable on the diapophyses of the fourth thoracic vertebra, and on the seventh assume the form of conical tubercles. On the eighth and following vertebræ they are flat, and are last distinguishable on the seventh caudal vertebra. Facets for the articulation of the tubercles of the ribs occur on the diapophyses of the first to the seventh thoracic vertebræ. On the latter vertebra the first transverse process appears as a short projection on the side of the centrum. On the eighth thoracic vertebra, the transverse process is broad and flat, with the anterior margin bent upward, and is about 48 mm. long. The base of the neural arch is strongly concave externally. The transverse process of the ninth thoracic vertebra is similar to the preceding one, but broader and not bent upward anteriorly. The base of the neural arch is also concave in this vertebra. The ends of the transverse processes of the eighth and ninth vertebræ are emarginate for the articulation of the ribs. A median inferior ridge is first distinguishable on the seventh thoracic vertebra.
As far as can be learned from the descriptions of Turner, Grieg, and others, the thoracic vertebræ of europæus do not present any marked differences from those of bidens.
The transverse processes of the lumbar vertebræ are short, broad, and flat, and somewhat curved forward. They are expanded and rounded at the free ends. The centra increase in length posteriorly, the last lumbar having the greatest length of any vertebra in the column. The neural spines increase in length from the first lumbar to the fourth, those on the remaining lumbars being subequal, but the spine on the ninth lumbar is a little longer than the others. Median inferior ridges occur on all the lumbars and are strongest at the middle of the series. The height of the centrum of the ninth lumbar is 63 mm., width 73, and length 116. The highest neural spine is 233.
As above mentioned, the first of the vertebræ counted among the lumbars may be the last thoracic vertebra, but as there is no indication of an articular facet at the end of the transverse process it is not so considered in this place.
The lumbar vertebræ in M. bidens appears to be more nearly equal in length than in the present species, but are not different otherwise.
The spines of the caudal vertebræ decrease rapidly in height posteriorly, and disappear after the tenth caudal. The transverse processes resemble those of the lumbars, but are shorter. They are last distinguishable on the eighth caudal. The transverse process of the seventh caudal is perforated by a vertical foramen. Similar but much smaller foramina occur on the sides of the centra of the eighth and ninth caudals. In these vertebræ the inferior ridges are also pierced by foramina. In the fourth caudal a ridge appears on the side of the neural arch on a level with the top of the centrum, and similar ridges are found on the succeeding vertebræ as far as the ninth caudal. The last ten vertebræ are without processes or neural arches.
Sir William Turner states that the caudals of M. bidens are without vertical foramina, but the figure in Van Beneden and Gervais’ Osteography (plate 22) shows them in the same position as in M. europæus. The inferior ridges, however, appear to be imperforate in the former species.
The first seven pairs of ribs have both tubercle and head. The first is nearly as long as the second, and is very broad at the proximal end. In the seventh pair the head is double, one facet of the rib articulating with the facet on the posterior margin of the centrum of the sixth thoracic vertebra and the other with the short transverse process on the side of the centrum of the seventh thoracic vertebra. The eighth and ninth pairs of ribs articulate only with the transverse processes of the eighth and ninth thoracic vertebræ, respectively. The ninth pair of ribs, as already stated, is nearly or quite as long as the eighth, from which it seems probable that a tenth short pair was present originally. There is, however, no trace of a facet for the articulation of such a rib on the end of the transverse process of what appears to be the first lumbar vertebra.
The only difference between the ribs of M. europæus and those of M. bidens appears to be that the first pair is much longer proportionately in the former species.
The sternum presents no differences of importance from that of M. bidens figured by Grieg,[24] except that the fourth and fifth segments are anchylosed together, both laterally and transversely, and that the two sides are symmetrical. (Pl. 13, fig. 2.)
The scapula of M. europæus presents an entirely different appearance from that of M. bidens as figured in Van Beneden and Gervais’ Osteography (plate 22). In europæus the scapula is very high anteriorly, the anterior border is convex forward and the anterior crest convex backward, bounding an elongated elliptical area. The posterior margin is straight. The acromion is short, with convex margins at the base, beyond which it narrows suddenly and terminates in a straight, cylindrical process, which is strongly inclined upward. The coracoid is as long as the acromion, nearly straight and horizontal, but expanded at the end. (Pl. 13, figs. 3, 4.)
The phalangeal formula of the Atlantic City specimen of M. europæus and those of three Norwegian specimens of M. bidens are as follows (the metacarpals being included):
Phalangeal formula of M. europæus and bidens.
| I. | II. | III. | IV. | V. | |
|---|---|---|---|---|---|
| M. europæus, Atlantic City: | |||||
| Left | 2 | 6 | 6 | 3+ | 3+ |
| Right | 2 | 7 | 6 | 4(+1?) | 4 |
| M. bidens: | |||||
| Landenæs | 1 | 6(5) | 5 | 4 | 3 |
| Fæø | 1 | 6 | 5 | 4 | 3 |
| Udsire | 1 | 6 | 6 | 5 | 4 |
In M. europæus the metacarpal of the third digit is much constricted in the middle. The shaft of the ulna is straight. Except in these particulars and the relatively small size of the whole pectoral limb, the latter appears not to differ materially from that of M. bidens. As shown above, the first digit in M. bidens consists of the metacarpal bone only, while in M. europæus a phalange is also present.
Dimensions of the skeleton of the Atlantic City specimen of M. europæus, No. 23846, U.S.N.M.
| mm. | |
|---|---|
| Length of the seven cervical vertebræa | 94 |
| Length of first, second, and third cervical vertebræa | 45 |
| Atlas: | |
| Greatest breadth | 156 |
| Greatest height | 103 |
| Height of neural canal | 36 |
| Greatest breadth across anterior articular facets | 96 |
| Axis, greatest breadth | 144 |
| Seventh cervical vertebra: | |
| Greatest breadth | 80 |
| Greatest height without inferior process | 117 |
| Greatest length of centrum | 14 |
| Greatest height of neural canal | 49 |
| First thoracic vertebra: | |
| Greatest height | 151 |
| Greatest breadth | 136 |
| Height of centrum | 37 |
| Length of centrum | 21 |
| Breadth of centrum (articular surface) | 48 |
| Height of neural spine | 61 |
| Height of neural canal | 53 |
| Seventh thoracic vertebra: | |
| Greatest height | 246 |
| Greatest breadth | 116 |
| Height of centrum | 35 |
| Length of centrum | 69 |
| Breadth of centrum | 46 |
| Breadth between transverse processes | 66 |
| Eighth thoracic vertebra: | |
| Greatest height | 246 |
| Greatest breadth (between transverse processes) | 142 |
| Height of centrum | 39 |
| Length of centrum | 73 |
| Breadth of centrum | 47 |
| First lumbar vertebra: | |
| Greatest height | 263 |
| Greatest breadth (between transverse processes) | 215 |
| Height of centrum (anterior) | 43 |
| Length of centrum | 83 |
| Breadth of centrum | 53 |
| First caudal vertebra: | |
| Greatest height | 263 |
| Greatest breadth (between transverse processes) | 207 |
| Height of centrum (anterior) | 65 |
| Length of centrum | 113 |
| Breadth of centrum | 67 |
| Seventh caudal vertebra: | |
| Greatest height | 153 |
| Greatest breadth | 87 |
| Height of centrum (without hypapophysis) | 66 |
| Length of centrum | 84 |
| Breadth of centrum | 70 |
| Length of last 10 caudal vertebræ | 285 |
| Sternum: | |
| Total length | 404 |
| Length of manubrium | 165 |
| Greatest breadth of manubrium | 134 |
| Depth of anterior notch of manubrium | 37 |
| Scapula: | |
| Length | 247 |
| Depth | 161 |
| Length of acromion | b44 |
| Length of coracoid | 59 |
| Humerus, length | 107 |
| Radius, length | 110 |
| Ulna, length | 100 |
| Pelvic bones, length | 51 |
Regarding the finding of the Atlantic City specimen and its exterior and gross anatomy, nothing has been published except brief references by Sir William Turner in 1889[25] and Dr. Glover M. Allen in 1906,[26] taken from a newspaper report of a communication made by myself before the Biological Society of Washington in 1889. On that account a somewhat detailed statement regarding it will be made in this place.
This individual (Pl. 41, figs. 1, 2) was a male, 12½ feet long. It was observed by the crew of life-saving station No. 28, near Atlantic City, New Jersey, on the afternoon of March 28, 1889. It had come inside the bar which skirts the coast at this point, and was apparently unable to find its way out. It was captured with some difficulty, after being wounded in the throat, and was dragged up on the beach near the station. Later in the day it was carried to the skating rink of Messrs. Johnson & McShea, at Atlantic City, where it was exhibited until Monday, April 1. On the next morning it was sent by express to Washington.
I examined it for the first time in Atlantic City on March 29. It was then lying on the floor of the skating rink in such a position that the under surfaces were concealed, and, as the teeth were not visible, I mistook it for a female. Upon its arrival in Washington, however, where it could be examined under more favorable circumstances, it proved to be a male. The following measurements were taken from the fresh specimen:
External dimensions of a specimen of M. europæus from Atlantic City, New Jersey.
| Ft. | in. | |
|---|---|---|
| Total length (in a straight line) | 12 | 6 |
| Tip of beak to base of dorsal fin (along the back) | 7 | 6½ |
| Tip of beak to base of pectoral fin (along the back) | 2 | 11 |
| Length of pectoral fin along center | 11 | |
| Greatest breadth of pectoral fin | 3¾ | |
| Height of dorsal fin (in a straight line) | 6 | |
| Length of base of dorsal fin | 1 | 2 |
| Breadth of flukes (tip to tip) | 2 | 11 |
| Depth of tail 14 inches in front of posterior margin of flukes | 8¼ | |
| Tip of beak to angle of mouth | 9¾ | |
| Tip of beak to eye | 1 | 8¼ |
| Length of eye | 1 | |
| Breadth of blowhole | 4 | |
| Tip of beak to right angle of blowhole | 1 | 6½ |
The general form was slender and elongate. The beak sloped gradually from its extremity to the forehead, and there was no constriction separating the beak from the remainder of the head. Behind the blowhole, the outline of the back commenced at a higher level, but immediately curved slightly downward, indicating the position of the neck. The line then rose gradually until the anterior base of the dorsal fin was reached. Behind the fin the outline sloped downward gradually to the flukes.
The dorsal fin was relatively small, falcate, and obtusely terminated. The distance in front of its anterior base was three-fifths of the total length. Its posterior margin was continuous with the ridge of the back, which extended to the flukes and terminated abruptly a little anterior to the middle point of the antero-posterior breadth of the flukes. In front of the fin the back was rounded.
The pectoral fins were small and were placed low down on the sides. Their anterior base was as far removed from the eye (in a straight line) as the eye was from the extremity of the beak. Their shape was somewhat different from that of the flippers of M. bidens figured by Sir William Turner.[27] Their anterior margin was nearly straight throughout; the extremity was evenly and distinctly rounded off. The posterior margin was slightly convex in the distal half and straight proximally.
The conformation of the region of the axilla was quite peculiar. The hard integument of the posterior margin of the flipper was continued proximally inward and forward to a point near the head of the humerus. The triangular area between this stiff edge and the side of the body was occupied by a thin, soft, wrinkled skin, in the middle of which the olecranon could be felt. On the side of the body this soft integument occupied an area nearly as large as the flipper, the underlying thick layer of blubber ending abruptly, especially below. A depression was thus formed in which the flippers could be placed so as to be almost in the same general plane with surrounding surfaces of the body. They are probably so placed when the animal is swimming.
The flukes had the general lunate form common to all species of the order. The posterior margin is not divided in the center. Its middle third was convex; its lateral thirds concave. In these and other respects the shape of the flukes agreed closely with Sir William Turner’s excellent figure of M. bidens.[28] The antero-posterior breadth of the flukes was, however, somewhat greater in proportion to their transverse breadth than is indicated in this figure. The caudal peduncle terminated above at a point 6½ inches in front of the posterior margin of the flukes. On this margin were situated three star-shaped white scars, which appeared to mark the points of attachment of crustacean parasites.
The margins of the upper jaw were very obtuse posteriorly, the rostrum being covered with a layer of blubber of gradually increasing thickness. A depression [22] bounded by gradually converging lines extended 4¼ inches back of the angle of the mouth.
The inferior surface of the bony palate extended below the level of the lips, and the sides of the former were visible upon looking into the mouth laterally.
The blowhole was large and somewhat unsymmetrically placed, the right angle being the more anterior. The concavity was forward.
The eye was situated a little below the line of the mouth and 20¼ inches from the extremity of the snout.
The external opening of the ear was 2⅞ inches behind the posterior angle of the eye, and a little below the line of the lower eyelid.
The two throat-furrows were of unequal length. The left furrow was 6¾ inches long, and its anterior end was distant 8⅝ inches from the extremity of the jaw. The right furrow did not extend quite so far forward, and was 7⅜ inches long.
The furrows converged posteriorly; they were separated by an interval of ⅝ inches anteriorly and 5⅛ inches posteriorly. Between the anterior ends of the main furrows was a small one, about an inch long, but it is doubtful whether this was a natural fissure. I did not observe it when the whale was in Atlantic City.
The natural color of the specimen had largely disappeared before I examined it, but Captain Gaskell and others who saw it while still fresh agreed that it was very dark slate-gray on the back, lighter on the sides, and whitish on the belly. I observed that a broad area between the pectoral fins was slate-gray, and contrasted with the white of the throat and belly. The whitish color ended somewhat abruptly and irregularly at the anus, and the flukes, as well as the pectoral and dorsal fins, were probably very dark slate-gray, or blackish, when fresh.
The epidermis was exceedingly smooth and glossy throughout.
The tongue was purplish-white. The roof of the mouth was black, except at the posterior end, where there was an irregular area of pinkish-white.
The integument of the roof of the mouth was smooth and shining. Its surface was convex at the extremity of the beak, but the central portion was concave, while at the posterior end it was again raised into a rounded pad. In these respects the shape of the integuments coincided with that of the underlying maxillæ, upon which they were closely fitted. The sides were rounded, and a shallow groove intervened between them and the lips. This groove was continued around the roof of the mouth behind, and formed a demarcation between this part and the œsophagus.
The tip of the tongue was 7½ inches from the extremity of the jaw. It was oval in outline, the extremity is obtuse, and it was entirely bound down. The margin was entire, and not crenulate, as in many dolphins.
Dorsal and ventral views of the stomach are shown in Pl. 40, figs. 1 and 2; a dorsal view of the lungs in Pl. 13, fig. 5; and of the perineum in Pl. 40, fig. 3. A description of the gross anatomy is reserved for a subsequent paper.
The external dimensions of the Atlantic City specimen of M. europæus are given in the following table, together with those of nine European specimens of M. bidens taken from various authors, and assembled here for purposes of comparison. The dimensions of the Annisquam specimen which, as already explained (p. 9), represents a third species, are also added.
External dimensions of Mesoplodon europæus, M. bidens, and M. densirostris.
| Measurements. | C | D | E | F | G | H | I | J | K | L | M |
|---|---|---|---|---|---|---|---|---|---|---|---|
| mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | |
| Total length | 3,810 | 4,877 | b4,828 | 4,597 | 4,572 | f4,315 | 4,605 | 3,870 | 3,700 | 3,450 | 3,708 |
| b12′6″ | 16′ | 16′2″ | 15′1″ | 15′± | =13′9″ | 15′1ࡩ″ | 12′8″ | 12′2″ | 11′4″ | 12′2″ | |
| Tip of upper jaw to blowhole | 470 | ... | ... | c572 | 559 | 602 | 475 | 530 | 500 | 440 | ... |
| Tip of upper jaw to eye | ... | ... | ... | ... | ... | ... | ... | ... | 520 | ... | ... |
| Tip of lower jaw to pectoral fin | d889 | ... | ... | ... | ... | 1,098 | 1,000 | 970 | 930 | 910 | ... |
| Back of dorsal to back of flukes | [1,156] | ... | e1,803 | ... | ... | [1,190] | 1,590 | 1,280 | [1,130] | 1,150 | ... |
| Length of base of dorsal fin | 356 | ... | 349 | 324 | ... | 366 | 400 | 340 | 330 | ... | ... |
| Length of eye | 25 | ... | ... | ... | ... | 46 | 40 | 37 | ... | ... | ... |
| Length of mouth (upper jaw) | 248 | 457 | 432 | ... | 343 | 373 | ... | ... | ... | ... | ... |
| Length of mouth (lower jaw) | ... | ... | 445 | 349 | 356 | 392 | ... | 320 | 320 | ... | 343 |
| Length of throat furrows | f173 | ... | 298 | ... | 254 | 248 | ... | ... | 300 | ... | ... |
| Distance between throat furrows posteriorly | 131 | ... | 241 | 229 | 178 | 157 | ... | ... | 217 | ... | 254± |
| Height of dorsal fin | 152 | ... | 191 | 203 | ... | 209 | 215 | 170 | 160 | 130 | ... |
| Breadth of flukes | 889 | ... | 1,118 | ... | ... | 994 | 1,130 | 1,000 | 820 | 680 | ... |
| Flukes to anus | ... | ... | ... | ... | ... | ... | 1,290 | 1,090 | [950] | 1,000 | 940 |
| Length of pectoral fink | g279 | ... | h546 | ... | ... | i392 | 515 | 440 | 380 | ... | ... |
| Greatest breadth of pectoral fin | 95 | ... | ... | ... | ... | 131 | 170 | 120 | 115 | ... | ... |
Since the foregoing account of europæus was written, a description of the type-skull, with two excellent photographic figures, has been published by L. Brasil,[29] of the Caën Museum. A comparison of the figures with those of the Atlantic City and Long Branch skulls on Pls. 2 and 8 of the present article, confirms the identification of the latter specimens with M. europæus. Besides a brief description of the type-skull M. Brasil’s paper contains measurements and two text figures of the right mandibular tooth, natural size.
This species was originally described from a single cranium of a young individual, which was collected by Dr. L. Stejneger on Bering Island, Commander Group, Bering Sea, in 1883. With but a single skull, the characters of the species could not be very satisfactorily defined, and some European cetologists have been inclined to doubt its validity.[30] In 1904, however, another skull was obtained by the National Museum, which made it certain that the species was entirely distinct from M. bidens or other known forms of the genus. Early in the year mentioned Dr. D. S. Jordan, president of Stanford University, called my attention to a small whale, which stranded on the coast of Oregon, 1½ miles south of the United States life-saving station on South Beach, Yaquina Bay, near Newport, in February, and proved later to represent the present species. Doctor Jordan’s information was obtained from Mr. J. G. Crawford, of Albany, Oregon, who wrote him in part as follows, under date of March 7, 1904:
Herewith I enclose a stereograph of a head of a member of the whale family, which I made at Yaquina Bay, Oregon. The animal was 17 feet long, with fluked tail, soft, smooth skin, blowhole on top of head, and two tusks in the mandible, but no [other] teeth in the mouth. The tusks are thin and apparently hollow. Length of head, 32 inches; width, 14 inches; height, 11 inches; blowhole, 5 inches. Eyes low on head. Width of mandible [jaw] at end: Upper, 1½ inches; lower, 1¾ inches. Width between tusks, 3 inches. The blubber was about 2 inches thick on the head. It went ashore about the 15th of February, 1½ miles south of the life-saving station on South Beach, 2½ miles south of Newport, Oregon. The head had been severed before I arrived.
A clipping from the Oregonian newspaper contains the following:
Albany, Oregon, March 2 [1904]. A peculiar specimen of the whale variety has been reported on the Oregon coast, near Newport. J. G. Crawford, of Albany, has just returned from a trip to Newport, where he made a picture of the head of the strange animal. The body was washed upon the beach during the recent storm which swept the coast. It is about 15 feet long. * * * Residents of the vicinity say they have never seen anything like it on the Oregon coast. * * * On either side of the mouth are two villainous-looking tusks several inches in length. They are at the back of the mouth, and extend up to a level with the top of the upper jaw. They are very wide and flat, squared on top. The mouth has no other teeth. * * *
[25]The head is equipped with a blowhole, like that of a whale. The eyes are very low, almost underneath the lower jaws.
The body is in a good state of preservation, the flesh having been torn but little by the birds.
On receipt of the foregoing information, letters were immediately addressed to Mr. Crawford and also to the keeper of the life-saving station at South Beach, Capt. Otto Wellander, asking that, if possible, the entire skeleton be preserved. Captain Wellander replied that the whale had not been dead long when washed ashore; that he had tried to find the body, but that the high tides had either carried it away or buried it under driftwood.
The skull when cleaned passed into the possession of Mr. J. G. Crawford, who sent to the Museum some excellent photographs of it, and also of the head before the flesh had been removed. Later he sent the skull itself to the Museum for my examination, and finally very generously presented it to the Museum in exchange.
The skull is that of an adult individual, in nearly perfect condition, with the mandible and teeth. The parts missing are the left malar, the left tympanic bone, the distal ends of the pterygoids and the proximal ends of the premaxillæ. (Pl. 3, fig. 2.)
The Oregon skull exhibits all the characters included in the original diagnosis of the species,[31] but two of these, namely, the lack of a groove in front of the premaxillary foramen, and the vertical position of the premaxillæ distally, I do not at present consider of any importance, as they are shared by M. bidens. The species, as represented by the Oregon skull, however, presents other characters which clearly differentiate it from any other species of the genius. As it is without a basirostral groove, it allies itself in that respect to M. bidens, europæus, and hectori. Unlike those species, it has the premaxillary foramen behind the maxillary foramen, and in this respect resembles densirostris and grayi. Perhaps the most salient characters in which stejnegeri differs from bidens and all other known species are the erect position and flat surface of the supraoccipital and the very prominent backward extension of the frontal plate of the maxilla. This backward extension is so great that when the beak is horizontal a vertical line through the posterior margin of the maxilla passes considerably behind the temporal fossa. The only species which approaches stejnegeri in this respect is hectori, but in the latter the supraoccipital instead of being flat above the condyles is very strongly convex.
Another very marked character of stejnegeri is that the extension of the lateral free margin of the orbital plate of the frontal, anterior to the orbit, is equal to the length of the orbit itself. In bidens and all other known species this extension is only from one-third to one-half the length of the orbit. Numerous other distinguishing characters will be mentioned in the course of the following description of stejnegeri, which is drawn from the adult Oregon skull, but modified when necessary by reference to the type skull from Bering Island. Comparisons are made chiefly with M. bidens, which is on the whole the best known species.
In the Oregon skull of stejnegeri, the breadth between the post-orbital processes does not exceed the length from the occipital condyles to the maxillary notches. The skull is, therefore, narrower in proportion to its length than in any other species of the genus except hectori, as represented by the skull figured by Flower. This skull was, however, that of a young individual. It is probable that in adults of this species the skull is broader than in stejnegeri.
In the latter species, again, the length of the brain-case, between the occipital condyles and the maxillary notches, is just equal to the distance from the latter point to the distal end of the maxillæ, and the rostrum, including the premaxillæ, is much shorter than in other species of Mesoplodon, except hectori, as represented by the young skull above mentioned.
The foramen magnum is very small, being less in width than the condyle on either side of it. In this respect it differs widely from bidens and other species (as far as can be ascertained from the figures available), except europæus, in which the relative size is about the same.
The supraoccipital rises vertically above each condyle to the very top of the skull, being neither convex nor strongly bent forward as in other species, and especially bidens. In the median line, however, while the occipital bone is flat immediately above the foramen magnum, it is deeply concave higher up and without a median ridge. The outline of the occipital crest, viewed from behind, is semicircular. In all the foregoing characters the occipital region differs widely from that of bidens and other species. The only close resemblance is found in the old skull of europæus from Long Branch, New Jersey, and even here the sides of the occipital above are far less prominent, their outline is much more convex, the occipital crest is angular, and the median depression is less pronounced.
Dorsal aspect (Pl. 3, figs. 1, 2).—The most noticeable feature of the upper surface of the skull is the large backward extension of the frontal plates of the maxillæ, the free margins of which converge strongly. The outline of the anteorbital region is rounded. The anteorbital notch is a shallow emargination. Anterior to this is a second still shallower emargination, the “pseudo-notch.” The margin between the two is much thickened, but does not form a distinct projection or tubercle, as in bidens and other species. The superior orifices of the nares are unsymmetrical as regards position, the left being somewhat in advance of the right. The maxillæ are concave around the maxillary foramen, and external to this foramen is an elongated ridge about as in europæus. The rostral portion of the maxillæ is broad at the base but tapers more rapidly than in bidens. The margin is thick. At the middle of the beak the outline of the maxillæ at a lower level is visible from above, which is not the case in bidens or europæus. The rostral portion of the premaxillæ is oblique proximally and vertical distally. Unlike bidens, these edges are sharp throughout. The mesethmoid ends opposite the maxillary foramina. Anterior to it is seen the concave upper surface of the vomer, which, however, becomes flat distally. At about the middle of the beak the anterior end is clasped by the posterior forked end of a “mesirostral” ossification, which has a convex surface. This ossification begins proximally below the edges of the premaxillæ, but its surface rises gradually anteriorly, and at the end of the beak it is much above [27] the premaxillæ. The end of the beak consists of the consolidated mass of the premaxillæ and mesirostral ossification, the whole being convex above and below, but flat on the sides. The ossification has a deep median groove, which reaches to within 95 mm. of the tip of the beak.
It will be seen that the conformation of the upper surface of the beak is quite different from that of bidens or any other species.
The maxillary foramina are large and directed forward, and have a distinct broad channel in front of them. In the Oregon skull the right foramen is single, but the left divided into two. The premaxillary foramina are a little behind the maxillary foramina. The distance between the maxillary foramina is less than that from the median line to the anteorbital notch. In bidens it is much greater.
Lateral aspect (Pl. 9, figs. 1, 2).—A most noteworthy feature of the skull when viewed from the side is the great length between the orbit and the maxillary notch, which far exceeds that found in bidens and other species, being equal to the length of the orbit itself. The latter is about as long as the temporal fossa, which is somewhat flattened above, as in europæus. The outline of the supraoccipital is straight and nearly vertical. The zygomatic is more massive even than europæus and is especially thick below. The inferior outline of the beak is convex proximally as in europæus and layardi. There is no basirostral groove, the edges of the maxillæ being very thick in front of the maxillary notch. Over the orbit the maxillæ are thick and beveled, but not raised as in bowdoini.
Ventral aspect (Pl. 6, figs. 1, 2).—The beak is convex in the proximal half, much as in europæus, but farther forward is concave, except in the median line, where there is a narrow ridge formed proximally by the vomer, which in the type skull appears as a narrow lozenge 60 mm. long. In the adult Oregon skull it is anchylosed with the premaxillæ. The maxillæ extend to within 107 mm. of the end of the beak. The under surface of the beak is much more like that of europæus than of bidens.
A narrow strip of the palatines extends around the base of the pterygoids in front, but the two strips do not meet in the median line. In the type-skull they do not extend inside the pterygoids. The expanded anterior end of the malar is very long and also forms the bottom of the maxillary notch, which is the case in europæus but not in bidens. The inferior borders of the pterygoids are convex anteriorly, as in europæus, and are continued laterally, so that the sinus is deep as in that species. The lachrymal is very long, the free margin having a length of 55 mm. The posterior margin of the zygomatic process is concave, rather than convex as in bidens.
The tympanic bulla does not differ materially from that of bidens in size or shape, as far as can be judged from the figures given in Van Beneden and Gervais’ Osteography (plate 26, figs. 4, 4a). The periotic is similar in size to the same bone in bidens, but the posterior end is more narrowly pointed and the anterior end is much lower, relatively. In europæus, as far as can be determined from the material at hand, the form and size of the earbone is similar to that of stejnegeri, but in the latter the anterior margin of the tympanic bulla is more nearly transverse and the posterior inferior groove is curved. (Pl. 35, fig. 2.)
In the Annisquam skull, supposed to represent densirostris, although from a young individual, the earbone is very much larger, especially the periotic, which is also quite differently shaped.
The mandible of stejnegeri is much broken in the region of the angle on both sides, but otherwise complete. As compared with a mandible of an adult bidens, the most conspicuous differences are the shortness of the symphysis, the sharp upward bend of the inferior margin anteriorly, and the large size of the alveolus. The symphysis in the adult Oregon specimen of stejnegeri is 140 mm. long, or scarcely more than in the young specimen of europæus from New Jersey, and exactly the same as in the adult type-specimen of the latter species, as figured by Van Beneden and Gervais. The alveolus lies entirely behind the symphysis, its anterior end being 160 mm. from the anterior end of the jaw. It is 113 mm. long and 18 mm. wide. The mandible is 62 mm. high at its middle point. The coronoid process is more anteriorly situated than in bidens and the portion of the posterior margin of the ramus which remains indicates that the angle was strongly directed backward. (Pl. 11, fig. 4; pl. 12, fig. 1.)
The teeth are remarkable for their size and form. They are somewhat more than twice as broad as teeth of adult males of bidens, as shown by the figures of Lankester[32] and Grieg,[33] and also a little longer. They are, in fact, probably broader than, or at least as broad as, the teeth of any other species of Mesoplodon, not excepting layardi. Sir William Turner remarks regarding a specimen of layardi examined by him that “the breadth of the tooth, where it emerged from the alveolus, was 3½ inches.”[34] He does not state, however, whether the measurement was taken along the top of the alveolus, at an angle with the transverse axis of the tooth, or along the transverse axis itself. At all events, the teeth figured by Owen and others are much less than 3½ inches broad. The teeth of adult europæus are only 2 inches broad, and of bidens, as already stated, 1½ inches broad.
In stejnegeri (Pl. 12, figs. 1-3) the portion of the tooth above the alveolus is inclined slightly inward and backward, but the pointed tip curves outward so as to be vertical. When extracted from the alveolus, the whole tooth is found to be concave internally and convex externally. The posterior margin is convex and the anterior sinuous, a slight convexity occurring on the portion which projects above the alveolus. In this place the outer coating of cement is broken through, showing the underlying dentine or osteo-dentine, which is somewhat corroded or absorbed. This is particularly noticeable on the left tooth.
The upper margin of the tooth is transverse, or nearly at right angles with the anterior and posterior margins. The posterior angle is rounded and the anterior raised into an acute point by the projection of the dentine as a distinct, sharp cusp. [29] The inferior end of the tooth is cut off obliquely and the margin is broken by numerous prominent rugosities. The surface of all that part of the tooth which is contained in the alveolus and covered by the gum above it is rugose, while the part above the gum is quite smooth and highly polished.
The right tooth has the following dimensions (in straight lines): Length of anterior border, 150 mm.; length of posterior border, 107; length of superior border, 54; length of inferior border, 86; average length of exposed dentine tip, 10; greatest breadth of tooth, antero-posteriorly, 81; greatest breadth of tooth, transversely, 15; distance from center of base of exposed portion, when in position in the alveolus, to tip of dentine projection, 82; distance from center of base of portion above the gum to tip of dentine projection, 70; distance from center of base of portion above the gum to center of inferior margin, 76.
The dimensions of the skulls are as follows, those of the type-specimen having been revised and corrected:
Dimensions of two skulls of M. stejnegeri.
| Measurements. | A | B |
|---|---|---|
| Total length | 715 | a633 |
| Length of rostrum | 413 | a325 |
| Distance from occipital condyles to distal end of maxillæ | 612 | 567 |
| Breadth between centers of orbits | 309 | 279 |
| Breadth between zygomatic processes | 310 | 278 |
| Breadth between temporal fossæ | 228 | 212 |
| Breadth between postorbital processes of frontals | 323 | ... |
| Breadth of rostrum at base (between maxillary notches) | 172 | b158 |
| Breadth of rostrum at middle | 40 | 44 |
| Depth of rostrum at middle | 52 | 42+ |
| Greatest breadth of anterior nares | 56 | 54 |
| Greatest breadth of premaxillæ proximally | 130 | 118 |
| Greatest breadth of premaxillæ in front of nares | 108 | 109 |
| Length of temporal fossa | 92 | 86 |
| Depth of temporal fossa | 63 | 46 |
| Antero-posterior length of orbit | 96 | 82 |
| Breadth of foramen magnum | 38 | 39 |
| Length of tympanic bulla | 48 | ... |
| Breadth of tympanic bulla | 32 | ... |
| Length of mandible | 610 | ... |
| Length of symphysis | 138 | ... |
| Distance from anterior end of mandible to alveolus | 166 | ... |
The photograph of the head (Pl. 40, fig. 4) shows that the end of the beak was quite blunt, and the lower jaw quite a little longer than the upper. The superior margin of the lower jaw, which is concave in front of the tooth, is strongly convex and elevated at the side of it and behind it. The inferior margin of the upper jaw [30] is straight anteriorly, but farther back appears to be pressed upward by the tooth. An examination of the skull shows that the mandible can be lowered so that the teeth are below the upper jaw, but when so lowered the space between the teeth and the upper jaw on each side is barely a quarter of an inch (6 mm.). With the integuments in place, it is doubtful whether the mouth could be opened any wider than is shown in the photograph. The convexity of the head, shape of the blowhole, position of the eye, etc., do not appear to differ materially from the same characters in adults of M. bidens.
It has not seemed to me necessary in the present connection to attempt to cite all the multitudinous names which have been given to this species, especially as those zoologists most competent to judge, including Van Beneden, Flower, and Turner, after detailed consideration, have concluded that but one species of Ziphius, or at most two species, exist at present.[35]
Nearly all the skulls in European museums are assigned by the zoologists mentioned to Z. cavirostris proper, but some doubt has been entertained regarding two or three European skulls, and one specimen from Argentina, described by Burmeister. These last-mentioned specimens have been thought to possibly represent a second species, Z. gervaisii. The principal characters of the latter are the narrow, flat premaxillæ, the lack of a prominent mesirostral ossification, and small teeth. From the large series of skulls in the National Museum, I am able to dispose of the doubt concerning Z. gervaisii. I find that wherever the characters above mentioned occur the sex (when known) is female. There is every reason, therefore, to believe that Z. gervaisii is the female of Z. cavirostris.[36] I will return to this point again later.
In 1865 Cope described a species from Charleston, South Carolina, under the name of Hyperoödon semijunctus. In 1886 I referred it to the genus Ziphius, but [31] was in doubt as to its specific identity. I thought that it might represent Z. gervaisii, which is interesting in the present connection because the type-specimen was a female.
In 1883 Dr. L. Stejneger described a species which he had discovered on Bering Island, Bering Sea, under the name of Z. grebnitzkii. Through the instrumentality of Doctor Stejneger and Governor Grebnitzki, the National Museum later received a large series of skulls from the same locality. The question of whether this species is identical with Z. cavirostris, or distinct, has caused me much study, and forms the principal subject of this chapter.
The National Museum has at present the following material, which may be considered as certainly representing Z. cavirostris:
1. A complete skeleton and cast of an adult female, 19 feet 4 inches long, obtained at Barnegat City, New Jersey, October 3, 1883. Cat. No. 20971.
2. A complete skeleton and photographs of an adult male, 20 feet 1 inch long, obtained at Newport, Rhode Island, in 1901, through Dr. E. A. Mearns, Mr. L. di Z. Mearns, and Capt. Gus Soderman. Cat. No. 49599.
3. The collection contains also the skeleton of the young female individual obtained at Charleston, South Carolina, prior to 1865, which constitutes the type of Hyperoödon semijunctus Cope. It was originally in the Charleston College Museum, but later was received by the National Museum in exchange. This individual was between 12 and 13 feet long. Cat. No. 21975.
In addition, the national collections contain the following material, known to, or supposed to, represent the species Z. grebnitzkii:
4. Cat. No. 20993. Skull of a male (?).[37] Collected by Dr. L. Stejneger in Bering Island, 1882. Orig. No. 1521. Type of Ziphius grebnitzkii.
5. Cat. No. 21245. Skull. Orig. No. 1758.
6. Cat. No. 21246. Skull. Orig. No. 2531.
7. Cat. No. 21247. Skull. Orig. No. 1849.
8. Cat. No. 21248. Skull of a male (?).
9. Cat. No. 83991. Skull.
The five skulls preceding were also collected by Doctor Stejneger in Bering Island in 1882 and 1883.
10. Cat. No. 22069. Skull of a female (?).[37]
11. Cat. No. 22874. Skull.
12. Cat. No. 22875. Bones of an immature individual.
These three specimens were collected and presented by N. Grebnitzki.
13. Cat. No. 142579. A series of photographs of an individual captured in Kiska Harbor, Alaska, September, 1904. Presented by Dr. J. Hobart Egbert.
14. Cat. No. 84906. Photograph of the skeleton of an individual washed ashore at St. Simon Island, Georgia, in 1893, and belonging to Mr. W. Arnold.
In the genus Ziphius, as in other ziphioid genera, a study of the characters of the skull appears to afford the best basis for discrimination of species. We have first to consider whether the North American species is the same as the European and New [32] Zealand species, and afterwards whether the North Pacific species is identical with or distinct from these.
The published measurements of specimens from the coasts of Europe and New Zealand, currently believed to represent the single species Z. cavirostris, are rather meager, and, furthermore, prove, on examination, to present so little uniformity that they are of limited use for comparison with measurements of skulls from the Atlantic coast of the United States. About all that can be said is that the latter skulls are of about the same size as the former and that the proportions do not present any striking differences. For detailed measurements of the American skulls, see page 53.
On account of the uncertainty as regards the measurements, I have had recourse to the published descriptions and figures, especially those of Van Beneden, Sir William Turner, and Doctor Haast. So far as I can perceive, there is nothing in these descriptions that is not applicable to the skulls Nos. 49599 and 20971, from Newport, Rhode Island, and Barnegat City, New Jersey, respectively, in the National Museum, and I can find no reason for regarding the latter other than as representatives of Z. cavirostris.
Of the Newport specimen, No. 49599, the Museum has the complete skeleton, together with external measurements and a photograph. From data at hand it appears that the animal was originally obtained in Narragansett Bay about October 30, 1901, and afterwards towed to Fort Adams, near Newport. A few days later it was sent adrift again and stranded in the harbor of Dutch Island, near Canonicut Island, which is opposite Newport. While at Fort Adams its existence was made known to the Museum by Dr. E. A. Mearns, U. S. Army, and his son, Louis Mearns; and a preparator was sent to obtain the skeleton. With the aid of Captain Soderman, of the government tug Monroe, he found it at Dutch Island, and reported that it was a male, 20 feet 1 inch in length, measured along the curves of the back (18 feet 6 inches in a straight line). The epidermis was nearly all lacking, but the back appeared to have been black. The length in a straight line, as reported by Mr. Louis Mearns, was 19 feet. The complete measurements taken by the preparator, Mr. J. W. Scollick, are as follows:
External dimensions of Ziphius cavirostris, male, Cat. No. 49599, U.S.N.M., Newport, Rhode Island.
| Ft. | in. | |
|---|---|---|
| Total length, along curve of back | 20 | 1 |
| Total length, in straight line | 18 | 6 |
| Tip of snout to posterior margin of dorsal fin | 13 | 10 |
| Tip of snout to axilla | 5 | 2 |
| Tip of snout to eye | 2 | 5½ |
| Tip of snout to anterior margin of blowhole | 2 | 4 |
| Length of mouth | 1 | 1 |
| Breadth of blowhole | 0 | 5½ |
| Length of pectoral fin, from head of humerus to tip, straight | 2 | 2 |
| Vertical height of dorsal fin | 0 | 10 |
| Breadth of flukes, from tip to tip | 5 | 3 |
| Greatest girth (estimated) | 10 | 0 |
The breadth of the pectoral fin, as shown by the skeleton, was 5¾ inches.
The photograph, which is reproduced in Pl. 41, fig. 4, gives a good idea of the general form of the animal.
Of the Barnegat City specimen, No. 20971, the Museum has the complete skeleton, together with a cast of one-half of the entire animal, and another of the head, and some measurements, all of which were obtained by Mr. William Palmer and myself October 3, 1883. The Museum received notice of the stranding of this specimen from Capt. J. H. Ridgway, of the United States life-saving station at Barnegat City. It was an adult female, 19 feet 4 inches long in a straight line. The complete measurements, taken in straight lines with a rod and cord, are as follows:
External dimensions of Ziphius cavirostris, female, Cat. No. 20971, U.S.N.M., Barnegat City, New Jersey.
(Measured in straight lines with rope and bar.)
| Ft. | in. | |
|---|---|---|
| Total length | 19 | 4 |
| Tip of snout to eyes | 2 | 1 |
| Tip of snout to blowhole | 2 | 0 |
| Tip of snout to anterior base of pectoral fin | 3 | 10½ |
| Tip of snout to anterior base of dorsal fin | 12 | 0 |
| Tip of snout to anterior angle of vent | 12 | 3½ |
| Tip of snout to corner of mouth | 11½ | |
| Length of anterior margin of pectoral fin | 2 | 1½ |
| Length along center of pectoral fin | 1 | 7 |
| Greatest breadth across pectoral fin | 6¾ | |
| Length of anterior margin of dorsal fin | 1 | 6 |
| Length of base of dorsal fin | 1 | 0 |
| Vertical height of dorsal fin | 1 | 0 |
| Breadth of flukes from tip to tip | 5 | 5 |
| Antero-posterior length of flukes | 1 | 7 |
| Length of eye | 2 | |
| Breadth of eye | 1 | |
| Girth around eyes | 3 | 1½ |
| Girth at anterior margin of dorsal fin | 7 | 0½ |
| Girth at root of pectoral fins | 6 | 0½ |
| Breadth of lower jaw at middle of length | 4½ | |
| Breadth of upper jaw at middle of length | 5 | |
| Breadth of blowhole | 5 | |
| Distance from posterior angle of eye to ear | 4½ |
I neglected to make a full description of the color, but noted that it was stone gray, lighter above and darker below; snout nearly white. The cast, which was painted from a sketch made at Barnegat City and from pieces of skin brought to Washington, bears out this note in general, but with modifications. The color of the body as a whole is gray tinged with dull yellowish. The gray is darker on the back than on the belly, but on the latter is a large area of dark brown, reaching from near the pectoral fins to and beyond the anus, and halfway up on the sides. On this dark area are several large oval whitish blotches, some two inches in [34] diameter. Both upper and lower jaws nearly to the angle of the mouth are cream white. On the sides and belly the gray color is speckled with black spots of about the size of a grain of wheat. The pectoral fins are dark gray above and below; the flukes were similarly colored.
A comparison of the dimensions of the two specimens above described with those of European and New Zealand specimens is afforded by the following table (the measurements being reduced to percentages of the total length):
External dimensions of Ziphius cavirostris. (Reduced to percentages of the total length.)
| Measurements. | Aa | Ab | Bb | C | D | E | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ft. | in. | Ft. | in. | Ft. | in. | Ft. | in. | Ft. | in. | Ft. | in. | |
| Total length | 20 | 1 | 18 | 6 | 19 | 4 | 19 | 6 | 19 | 0 | 12 | 11½ |
| Per cent. | Per cent. | Per cent. | Per cent. | Per cent. | Per cent. | |||||||
| Tip of snout to posterior margin of dorsal | 69.0 | 74.8 | 67.2 | [67.1] | [78.5] | [70.8] | ||||||
| Tip of snout to axilla | 25.3 | 28.0 | c20.0 | d24.4 | ... | c[25.0] | ||||||
| Tip of snout to eye | k12.2 | 13.3 | 10.8 | 12.8 | ... | 10.9 | ||||||
| Tip of snout to anterior end blowhole | 11.2 | 12.6 | 10.4 | ... | ... | 11.4 | ||||||
| Length of mouth | 5.4 | 5.9 | e 5.0 | 6.4 | ... | [ f] 5.3 | ||||||
| Breadth of blowhole | 2.3 | 2.5 | 2.2 | 2.6 | ... | 1.2 | ||||||
| Length of pectoral from head of humerus | 10.8 | 11.7 | ... | g12.8 | ... | ... | ||||||
| Length of pectoral from axilla | h7.5 | 8.1 | hi8.1 | ... | g 8.3 | 8.6 | ||||||
| Greatest breadth of pectoral fin | j2.4 | j2.6 | 2.9 | 3.0 | 2.9 | 3.0 | ||||||
| Vertical height of dorsal fin | 4.1 | 4.5 | 5.2 | 3.4 | 3.5 | 4.3 | ||||||
| Breadth of flukes, tip to tip fin | 26.1 | 28.4 | 28.0 | 31.2 | ... | 27.3 | ||||||
The close correspondence in proportions shown in this table favors the idea of specific identity, and taken with the similarity in size, and characters of the skull, warrants, I think, the assumption that the specimens from the Atlantic coast of the United States belong to Z. cavirostris.
It should be remarked, however, that the Barnegat City specimen does not agree in color with any of the European or New Zealand specimens. On the other hand, the latter show a most extraordinary diversity in color, some being black, with the head and back as far as the dorsal fin white; others all black above, white [35] below, and the head black and brown. The color of the young specimen from Buenos Ayres, Argentina, is described by Burmeister as follows:
All the body of the animal is of a light gray color, a little yellowish, resembling the color of light ash, but much darker on the back and much lighter on the belly. The fins are much darker than the back—almost black—and the large fin of the tail has a very pure white area of irregular shape on the underside.
If the indications from the skull and proportions are trustworthy Z. cavirostris must be a species in which the color is very variable, differing perhaps in the two sexes, or with differences in age. This is, however, by no means certain at present, and whether the diversities of color reported in different specimens are merely individual variations, or are due to post-mortem changes, remains to be discovered. It will be noticed that the color of the Argentine specimen is nearest to that of the Barnegat City specimen.
The type-specimen of Ziphius semijunctus (Cope), as already mentioned, is a young female.[38] The most noticeable characters which it presents are that the premaxillæ are flat proximally, and that the teeth are small, sharp-pointed and open at the roots. The form of the teeth is undoubtedly due to immaturity, but [36] as the shape of the premaxillæ is similar to that found in the nominal species gervaisii, it might be thought necessary to refer semijunctus to the latter species. As will be shown later, however, this form of the premaxillæ appears to be characteristic of the adult female of cavirostris, and of immature individuals of either sex, the young, as in many kinds of animals, resembling the adult female rather than the male.
I have been able to find but one character in the skull of semijunctus which might be regarded as specific. This is that the lachrymal bone is thick distally, and cut off square at the end. In other specimens of Ziphius examined it is thin and flat, and rounded or pointed at the end. As there is much individual variation in the form of the lachrymal, this peculiarity alone is, in my opinion, an insufficient indication of the validity of the species.
A comparison of the skeletons of the three individuals from the Atlantic coast of the United States reveals a number of differences of more or less importance. Were it not for the lack of reliable differences in the skulls, it might be considered that these variations in other parts of the skeletons indicated specific difference. I am disposed, however, since the Barnegat and Newport specimens are of opposite sexes, to regard them partly as sexual and partly as individual. In the case of the Charleston specimen (semijunctus), the skeleton, besides being immature, has been very much damaged by careless handling, and nearly all the bones are somewhat abraded. It is, therefore, only available to a limited extent for purposes of comparison. As no description of a Ziphius skeleton from the coast of the United States has, so far as I am aware, been published hitherto, and as descriptions of skeletons of Old World specimens are few and rather brief, I shall give below a detailed comparative description of the American specimens. For the sake of brevity, I shall refer to each specimen merely by the locality.
The vertebral formula in the three North American specimens and in four Old World specimens and Burmeister’s Argentine specimen is as follows:
Vertebral formula of Ziphius cavirostris.
| Locality and sex. | C. | Th. | L. | Ca. | Total. |
|---|---|---|---|---|---|
| Newport, Rhode Island, male | 7 | 9 | 10 | 20 | 46 |
| Barnegat City, New Jersey, female | 7 | 9 | 11 | 18(+1?) | 46(?) |
| Charleston, South Carolina, female | 7 | 10 | 10 | 16(+3?) | 46(?) |
| Holma, Sweden (Malm) | 7 | 10 | 10 | 18(+1) | 46 |
| Pisa Museum (Van Beneden) | 7 | 9 | 11 | 16+ | 43+ |
| Warrington, New Zealand (Scott and Parker) | 7 | 10 | 9 | 20 | 46 |
| Lyttleton Harbor, New Zealand (Haast) | 7 | 9 | 11 | 19 | 46 |
| Buenos Ayres, Argentina (Burmeister), male | 7 | 10 | 10 | 22 | 49 |
In the figures of the Argentine specimen the last ten caudals are practically without characters, and it is perhaps allowable to question whether the terminal two or three were not added to make an even taper to the end of the column. If such be not the case, this specimen had more vertebræ than any other.
Newport (male).—The seventh cervical vertebra presents a conical metapophysis, which on the first thoracic vertebra forms of a rather thick, long, declining process ending in a facet for the tubercle of the first rib. This metapophysis maintains nearly the same form as far as the sixth thoracic vertebra, but on the third thoracic a mammiliform process makes its appearance on the anterior margin near the tip, and becomes more prominent on each succeeding vertebra. On the seventh thoracic it becomes larger, thin, and upright, and widely separated from the articular facet for the tubercle of the rib. On the centrum of this vertebra lower down is a second much larger rugose articular facet. On the eighth thoracic vertebra the upper articular process disappears altogether and is replaced by a transverse process on a lower level, with a facet at the free end for the rib. On the ninth thoracic the transverse processes are larger and nearly straight. They are longer on the first lumbar and incline a little forward. Those of the succeeding vertebræ are similar, but decrease gradually in length, while somewhat increasing in breadth. They are last traceable on the ninth caudal. On the eighth caudal they are perforated by a foramen.
All the vertebræ from the first cervical backward have neural spines as far as and including the eleventh caudal. The spine on the first thoracic is rather short, narrow and pointed. These spines increase in height in succeeding vertebræ as far as the sixth lumbar; at the same time the breadth increases antero-posteriorly and the tip becomes expanded. The spines are nearly equally high on all the succeeding lumbars, but begin to decrease on the caudals and disappear altogether on the eleventh caudal.
The anterior zygapophyses and metapophyses maintain a nearly constant position close to the top of the centra throughout the column, from the seventh thoracic backward, and are vertical, thin, and oblong, squared or rounded. They begin to decrease in size noticeably on the first caudal, and on the seventh caudal are mere swellings at the sides of the nearly horizontal plate from which the neural spine springs. They are traceable as far as the twelfth caudal.
A ridge appears on the side of the neural arch near its base on the fifth caudal and is stronger and very marked on those following, to the ninth caudal. A ridge unites the anterior and posterior facets for the chevrons on the ninth and succeeding caudals.
Barnegat City (female).—Unlike the Newport skeleton, there are no neural spines on the fifth, sixth, and seventh cervicals. The spine on the first thoracic vertebra is quite short and sharp, and on the second, third, and fourth thoracics also is rather pointed, though of increased length. There is no metapophysis on the seventh cervical.
On the seventh thoracic the facet for the tubercle of the rib, instead of being very prominent, becomes inconspicuous. The metapophysis is flat and squared, and there is no lower facet on the side of the centrum. On the eighth thoracic the metapophysis is thin, squared, and vertical, and a well-formed transverse process appears on the side of the centrum. The transverse processes of the ninth thoracic are a little curved backward, and on the first lumbar and succeeding vertebræ bent forward. These processes are less tapering on all the lumbars than in the Newport skeleton. They disappear on the eighth caudal. None is perforated.
The longest neural spine is on the sixth lumbar, and on all the lumbars both the anterior and posterior edges are somewhat convex. Hence their shape is rather different from those of the Newport skeleton, in which the anterior margins are somewhat concave. The tips of the spines are rather suddenly expanded. The spines of the caudals are rather more expanded at the tip and more inclined backward than in the Newport skeleton. They disappear on the eleventh caudal.
The horizontal plate joining the metapophyses is noticeable on the fifth caudal. The ridge on the side of the neural arch is first noticeable on the fourth caudal and is very strong on the fifth, sixth, and seventh. The metapophyses are last traceable on the twelfth caudal.
Charleston (female, jr.).—This skeleton resembles the Newport one as regards the facets for the articulation of the tubercles of the ribs, except that the seventh thoracic resembles the sixth and has no lower facet on the side of the centrum. The transverse processes of the ninth thoracic are rather strongly curved backward, while those on the last thoracic and first lumbar are nearly straight. On succeeding vertebræ they are inclined forward. They are last traceable on the eighth or ninth caudal (vertebra 35 or 36). None is perforated by a foramen.
Though the vertebræ are defective, there appear to have been no neural spines on the fourth to the seventh cervicals, inclusive. The spine on the first thoracic is short, and on the first to the fourth is pointed. The spine disappears on the tenth caudal (vertebra 37).
The metapophyses assume the vertical position on the eighth thoracic. The last of these processes is barely traceable on the tenth caudal (vertebra 37). The ridge on the side of the neural arch is well marked on the fifth to the ninth caudals, inclusive. On the seventh caudal (vertebra 34) the anterior and posterior facets for the chevrons are united on the right side, and on the eighth caudal and succeeding vertebræ on both sides.
Barnegat City (female).—The first four cervicals are united. The foramen above the anterior articular facets of the atlas is complete, and the edges of these facets are raised. The inferior lateral process is flat, broad, and strongly bent backward.
Second cervical: Inferior lateral process nearly as long as that of the first cervical; broad, flat, and bent backward parallel with the process of the first cervical. Superior lateral process short, strong, and flat. A large incomplete foramen between it and the inferior process.
Third cervical: A short, conical inferior process, curved forward.
Fourth cervical: Similar, but with smaller and shorter inferior process. Neural arch and spine complete; the latter fused with the preceding spines. Arch not reducing the size of the neural canal.
Fifth cervical: Arch and spine broken. Arch nearly as broad as the anterior epiphysis of the centrum. Inferior lateral process short, straight, and directed obliquely outward.
Sixth cervical: Spine broken. Arch complete, nearly as wide as the anterior epiphysis. Inferior lateral process short, thick, knobbed, and directed obliquely outward and a very little forward. The left longer.
Seventh cervical: Spine obsolete. Arch complete, as wide as the anterior epiphysis. No superior lateral process or metapophysis. A thick articular facet for the head of the first rib on the middle of the side of the centrum. No inferior lateral process.
Fused spines of the first to fourth cervicals bent backward; the mass broad antero-posteriorly and rounded at the tip.
Newport (male).—First cervical with the foramen over the anterior articular facets incomplete, and the borders of the facets less raised. The facets also broader and more declined. Inferior lateral process thicker, somewhat tapering, and nearly transverse.
Second cervical: Inferior lateral process much shorter than that of first cervical, about parallel with it, but with the tip bent forward. Superior lateral process short, thick, and bent backward; joined to the inferior process on the right side, inclosing an oval foramen.
Third cervical: A short, straight, triangular superior process on the right side; that on the left short and blunt. Inferior lateral process long, thick, club-shaped, and curved backward.
Fourth cervical: Inferior lateral process similar to the last in shape, but shorter, broad and flat, and only slightly curved backward. Neural arch and spine separate from those of the third cervical; the arch rather smaller than those preceding it, and reducing the size of the neural canal.
Fused spines of the first to third cervicals nearly vertical, rather high, and obtusely pointed.
Fifth cervical: Spine pointed and quite long. Arch complete. Inferior lateral process short, squared, flattened, and directed outward obliquely.
Sixth cervical: Spine about as long as on the fifth cervical. Arch much narrower than the anterior epiphysis. Inferior lateral process prominent, thick, somewhat compressed, and directed downward.
Seventh cervical: Spine as high as the arch, obtusely pointed. Arch complete, as wide as the anterior epiphysis. A strong conical superior lateral process, or metapophysis, on a broad base, directed forward. An articular raised facet on the side of the centrum, directed obliquely backward. No inferior lateral process.
Charleston (female, jr.).—The first to fourth cervicals resemble those of the Newport skeleton, but the fourth entirely separate. All the lateral processes undeveloped, or broken off, except the right inferior lateral process of the atlas, [40] which is like that of the Newport specimen. The anterior foramen of the atlas is incomplete, as in that specimen, and the spines of the conjoined vertebræ are vertical and pointed. (Pl. 25, fig. 1.)
Fifth cervical: Spine wanting. Arch complete. Inferior lateral process undeveloped, or abraded.
Sixth cervical: Spine and processes broken. Arch wide.
Seventh cervical: Similar to that of the Newport skeleton, but the spine obsolete or broken.
Barnegat City (female).—First thoracic: Spine vertical, pointed, about as high as arch and centrum together. A moderately long process with articular facet for tubercle of rib on side of neural arch; facet elliptical and directed a little downward and forward. A smaller facet for head of second rib on posterior upper edge of centrum.
Seventh thoracic: Metapophyses long, extending horizontally, straight superiorly. A small articular facet on the outer side near the base, directed downward; strongest on right side. A very small facet on posterior upper edge of centrum, scarcely noticeable on right side. Neural spine rather narrow at tip; superior margin straight.
Eighth thoracic: Metapophyses squared and thin. A distinct transverse process on side of centrum about half as broad as the centrum is long, and as long as centrum is broad; flattened, squared, and a little curved backward and upward. Articular facet for rib elliptical and directed obliquely backward. A broad, shallow groove across base of transverse process, the anterior edge of which is emarginate proximally. Neural spine as in seventh thoracic.
Ninth thoracic: Metapophyses squared. Transverse process similar to that of eighth thoracic, but equal to centrum in length, little narrowed at base, and directed outward; anterior edge convex, posterior concave; articular facet occupying the posterior half of the distal edge. A very shallow groove proximally.
Newport (male).—First thoracic: Neural spine a little curved backward and rounded at tip; much higher than length of arch and centrum together. Articular facets as in Barnegat skeleton.
Seventh thoracic: Metapophyses similar in shape to those of Barnegat skeleton but with a very distinct facet on side of arch, terminating a process about as long as the greatest diameter of the facet; surface of facet rugose. Below this process, on side of centrum, a very large, oval, sessile facet, reaching forward nearly to the anterior face of the centrum and upward to its superior edge. A very low, small swelling on the posterior superior edge of centrum, probably indicating the point of attachment of a cartilage connecting the head of the eighth rib. Neural spine expanded at free end, and superior margin rounded.
Eighth thoracic: Metapophyses similar to those of Barnegat skeleton. A distinct transverse process nearly as broad as the length of the centrum, oblong or squared, flat, directed somewhat backward, but not upward. Articular facet for rib not occupying whole of free end and only slightly directed backward; anterior margin as in Barnegat skeleton. Neural spine similar to that of seventh thoracic.
Ninth thoracic: Similar to that of Barnegat skeleton, but transverse process longer than centrum and directed a little downward, articular facet occupying less than posterior half of free margin; proximal groove inconspicuous; anterior and posterior margins nearly straight.
Charleston (female, jr.).—The centra of the thoracic, as well as the lumbar, vertebræ in this individual present inferior median keels, and more or less concave sides, which is not the case in the Barnegat and Newport skeletons. This can not be due to immaturity, as in a still younger individual, supposed to represent Ziphius grebnitzkii, the thoracic vertebræ are rounded below. The neural spines of the thoracic vertebræ are much less inclined backward in semijunctus than in the Newport and Barnegat skeletons, but this is doubtless connected with age, as the younger series of vertebræ already mentioned exhibits the peculiarity in a more marked degree. A similar modification dependent upon age appears to affect Hyperöodon, as will be seen by comparing Van Beneden and Gervais’ figures in the Osteography, plate 18.
First thoracic: Similar to that of Newport skeleton, but spine not higher than arch alone. (A little abraded at tip, but probably undeveloped.)
Seventh thoracic: Metapophyses short (abraded), incompletely developed. A distinct facet on side of same on an elongated process, as in Newport skeleton, but no second larger one on side of centrum. No facet on superior margin of centrum either anteriorly or posteriorly.
Eighth thoracic: Transverse process similar to that of Barnegat skeleton, but anterior edge nearly straight; process about one-half as broad as length of centrum. (Indications of immaturity.)
Barnegat City (female).—First lumbar: Similar to last thoracic, but transverse process expanded distally and slightly directed forward; a little longer than centrum; anterior and posterior edges emarginate proximally.
Eleventh lumbar (last): Centrum very long. Neural arch and spine very high, more than twice length of centrum. Spine inclined backward much beyond posterior face of centrum; anterior margin straight, posterior convex, tip expanded. Transverse process a little more than one-half length of centrum, somewhat expanded at distal end and curved forward so that tip is about in line with anterior face of centrum. Metapophyses close to centrum and to each other, semihexagonal in outline. A sharp median inferior ridge, and shallow posterior oblique channels on under side of centrum.
Newport (male).—First lumbar: Similar to that of Barnegat skeleton, but transverse processes considerably longer than the centrum and not expanded at tip; anterior edge straight, posterior only slightly emarginate proximally.
Tenth lumbar (last): Centrum like that in Barnegat skeleton. Neural arch and spine only slightly higher than length of centrum. Transverse process oblong, free margin nearly transverse; process inclined forward so that tip is a little beyond anterior face of centrum. Metapophyses close to centrum, rounded in outline. Neural spine much inclined backward; anterior edge concave, posterior convex, [42] tip expanded. A rounded inferior median ridge and very distinct oblique posterior channels on under side of centrum.
Charleston (female, jr.).—First lumbar: Similar to that of Barnegat skeleton, but transverse process directed outward and scarcely or not at all forward; length of process equal to that of centrum; tip rounded (due to immaturity).
Tenth lumbar (last): Centrum very long. Neural arch and spine a little less in height than length of centrum. Transverse process oblong, curved forward, more than one-half as long as centrum. Metapophyses similar to those of Newport skeleton. Inferior median ridge very sharp; lateral channels rather indistinct.
Barnegat City (female).—First caudal (vert. 28): Similar to last lumbar, but neural spine broader antero-posteriorly. Transverse process ࡪ length of centrum, inversely triangular, the tip much in advance of anterior face of centrum, free end somewhat rounded. Metapophyses similar to those of last lumbar. No median inferior ridge, but two short processes bearing facets for chevrons posteriorly and a very slight indication of similar process anteriorly, but without facets. Posterior inferior oblique channels indistinct.
Seventh caudal (vert. 34): Centrum (exclusive of chevron processes) nearly as deep as long. Neural arch and spine only a little higher than length of centrum, very much inclined backward and expanded at distal end; free border of spine straight. Metapophyses close to centrum, united nearly to tips by a horizontal plate. A ridge extends backward from their tips nearly across the arch. Another very prominent ridge traverses the centrum at the base of the arch. At the posterior end, a deep groove, convex forward, extends down the side of the centrum, making an emargination in the transverse process and proceeding thence down the lower side of centrum to its lower middle point, where it ends in a deep semicircular emargination between the anterior and posterior chevron facets. Transverse process a triangular stub, reaching nearly to the line of the anterior face of centrum. Chevron processes very large, and the median inferior surface of the centrum between them deeply grooved longitudinally.
Tenth caudal (vert. 37): Centrum as deep as long. Neural spine a low ridge, as long as the centrum, and extending beyond it posteriorly. No transverse processes. A foramen in side of centrum much above the middle and a similar one below. Close to the latter and below it another foramen pierces the ridge uniting the chevron processes, and appears below on side of longitudinal inferior median channel. Metapophyses small mammilliform processes on top of centrum.
Eleventh caudal (vert. 38): No processes. A very small neural spine. Posterior epiphysis strongly convex.
Twelfth caudal (vert. 39): A rounded mass without processes.
Thirteenth caudal (vert. 40): An oblong mass, with two grooves on each side, two widely separate foramina above and two closely approximated below, entering a common depression, with rounded projections on its borders.
Fourteenth caudal (vert. 41): Similar to thirteenth caudal, but with a single lateral groove.
Fifteenth caudal (vert. 42): Similar to fourteenth caudal, but sides extending upward and downward in a ridge. Inferior foramina nearly as far apart as superior and posterior epiphysis much smaller than anterior.
Sixteenth caudal (vert. 43): Similar to fifteenth caudal, but the disproportion of epiphyses greater and lateral ridges higher. Superior and inferior surfaces of centrum inclined.
Seventeenth caudal (vert. 44): Similar to preceding, but smaller.
Eighteenth caudal (vert. 45): Longer than high. Inferior ridge longer and larger than superior. Groove very large. Anterior face of centrum deeply concave, posterior flat. Posterior epiphysis very much smaller than anterior. Foramina very small, practically obliterated on right side.
Newport (male).—First caudal (vert. 27): Similar to last lumbar, but transverse process shorter, about two-thirds as long as centrum, oblong and but little constricted at base; distal margin nearly straight. The process does not extend forward quite to the line of the anterior face of centrum. No inferior median ridge, but strong posterior chevron processes. Postero-inferior oblique grooves very distinct.
Seventh caudal (vert. 33): Similar to the same vertebra in Barnegat skeleton, but neural spine more inclined backward and anterior border deeply concave. Metapophyses oblong, directed upward, not reaching anterior face of centrum as they do in Barnegat skeleton. Anterior face of centrum receding superiorly and the ridge opposite it on side of centrum shorter than in Barnegat skeleton. Ridge behind metapophyses indistinct. Postero-inferior oblique grooves as in Barnegat skeleton, but piercing transverse process, forming a foramen. Anterior and posterior chevron processes very large and receding very much, as do also the anterior and posterior faces of centrums.
Eleventh caudal (vert. 37): Similar to Barnegat skeleton, but spine shorter than centrum and not extending beyond it anteriorly or posteriorly. Metapophyses similar, but wider apart.
Twelfth caudal (vert. 38): Neural arch barely complete. No spine.
Thirteenth to nineteenth caudals (vert. 39-45): Similar to those of Barnegat skeleton.
Twentieth caudal (vert. 46): Rudely triangular, with a peg-like posterior projection, bearing the very small posterior epiphysis. No foramina. Anterior epiphysis deeply concave in middle.
Charleston (female, jr.).—First caudal (vert. 28): Similar to last lumbar, but only a faint inferior median ridge. Inferior outline of centrum antero-posteriorly very concave, which is not the case in the Barnegat and Newport skeletons. Posterior chevron processes prominent. Postero-inferior oblique grooves shallow.
Seventh caudal (vert. 34): Like the Newport skeleton. The transverse process not pierced or emarginate. Postero-inferior oblique grooves indistinct. Ridges on centrum very distinct. Right anterior and posterior chevron processes united and pierced by a foramen.
Tenth caudal (vert. 37): Similar to the same vertebra in Newport skeleton, but neural spine very short.
The number of chevrons in the North American and some other specimens is as follows:
| Newport, Rhode Island. | 11 |
| Barnegat City, New Jersey. | 8(+3?) |
| Charleston, South Carolina. | 8+ |
| Buenos Ayres, Argentina (Burmeister). | 11 |
| Holma, Sweden (Malm). | 9 |
| Littleton Harbor, New Zealand (Haast). | 10 |
| Warrington, New Zealand (Scott and Parker). | 9 |
| Pisa Museum (Van Beneden). | 9 |
The chevrons are similar in form in the three North American specimens, with some differences which will be pointed out below.
Newport (male).—The first chevron consists of a pair of bones which are not united. They are longer than deep, their depth indeed being less than that of any one of the succeeding bones except the tenth and eleventh. Each presents one strong superior articulating facet. Second chevron, elongated antero-posteriorly, but not much deeper than the first. Third chevron very deep and only equaled in that respect by the fourth; narrowed and rounded off below. Fourth chevron largest and broadest (antero-posteriorly) of the series; expanded below and the lower border transverse. Fifth to eighth similar in form, but less deep successively, and the lower border more rounded. Ninth similar to eighth, but smaller and thinner. Tenth similar to first, longer (antero-posteriorly) than deep. Eleventh similar to tenth in form, but smaller.
Barnegat City (female).—First chevron bone lacking. Second like that of Newport skeleton, but smaller. Third similar to second, but much larger and more produced posteriorly; quite unlike the third in the Newport skeleton in form, and much less deep. Fourth, largest and deepest of the series; anterior and posterior borders rounded, and the inferior border similar. Fifth to eighth similar in form, but successively less deep, and all more expanded below; inferior border nearly straight. Ninth similar to eighth, but depth not exceeding breadth; lower angles produced.
Charleston (female, jr.).—The chevrons of this specimen resemble those of the Newport skeleton, but on account of immaturity they are all more or less rounded. The two sides of the first chevron are united. The second is without the posterior angular projection seen in the other specimens. The third is the deepest of the series. The eighth is not deeper than long, and hence resembles the tenth chevron of the Newport skeleton in proportion, but is, of course, much smaller. Two or three chevrons are lacking from the posterior end of the series.
Barnegat City (female).—First rib shortest and broadest, but considerably broader at proximal end than at distal end. Head and tubercle close together. The succeeding ribs increase in length and decrease in breadth to the fifth or sixth. The third, fourth, and fifth are expanded and flattened at distal end. Seventh, eighth, and ninth successively shorter. Distance between head and tubercle [45] greater on second rib than on first, and on third is greater than on second. On the third to sixth, inclusive, the distance is about equal. The tubercle is scarcely distinguishable on the seventh rib, while on the eighth and ninth it is lacking, these ribs joining the transverse processes by a terminal facet only.
Newport (male).—Similar to those of the Barnegat skeleton, but first rib maintains nearly the same breadth throughout. Neck thicker than in Barnegat skeleton. Seventh rib terminates proximally in a single large rugose facet, which connects with a similar facet on side of centrum of seventh thoracic vertebra.
Charleston (female, jr.).—Similar to those of the Barnegat skeleton, but a distinct tubercle on the seventh rib. Eighth and ninth ribs end proximally in a transverse facet only, which is largest on the eighth. Tenth rib (represented by a fragment) only half as broad as the preceding ones and more nearly round in section.
Barnegat City (female).—Five segments. Manubrium wider than long, convex inferiorly. Deep anterior and posterior notches, about equal, the former with an angular projection on each side. Facet for cartilaginous sternal rib thick and prominent. Second segment wider than long, about equally notched anteriorly and posteriorly, the two sides anchylosed together by a bony bridge, about as wide as the notches are deep. Third and fourth segments similar to second but smaller; similarly notched; left portion a little longer than right. Fifth segment elongated, left side very much so; the two sides joined by a narrow bridge; posterior notch very deep.
Newport (male).—Similar to sternum of Barnegat skeleton, but manubrium scarcely wider than long; posterior notch much longer than anterior, with parallel sides. Second and third segments similar to those of Barnegat skeleton but sides of latter not completely anchylosed together. Fourth segment in two pieces, with a wide interval between. Fifth segment triangular with deep anterior, triangular notch, a narrow bridge, and short posterior prolongation (the left longer than the right).
Charleston (female, jr.).—Resembles the sternum of the Barnegat skeleton rather than that of Newport skeleton, but anterior parts cartilaginous. Opposite sides of second, third, and fifth segments anchylosed together and those of fourth segment nearly so. (Pl. 25, fig. 2).
Barnegat City (female).—Superior border irregular. Posterior angle acute. Anterior and posterior borders nearly straight. Ridges distinct. Acromion broad both at base and at tip, sharply bent upward, so as to be parallel with anterior border of scapula. Coracoid nearly as long as acromion, slender, a little curved upward, irregular and somewhat expanded at the end.
Newport (male).—Superior border irregularly rounded. Posterior angle obtuse, anterior angle projecting. Ridges indistinct. Anterior and posterior borders nearly straight, but irregular. Acromion broad at base, tapering toward the [46] tip, which is again somewhat expanded; bent upward, but not sufficiently to be parallel with anterior margin of blade. Coracoid rather thick, irregular, strongly expanded at tip.
Charleston (female, jr.).—Rather too much abraded for comparisons, but posterior margin more concave than in either of the other skeletons.
Barnegat City (female).—Fore limb much shorter than in the Newport skeleton. Humerus: Head quite oblique, the lower edge overhanging the shaft considerably on the ulnar side. Tuberosity level with upper surface of head, elliptical in outline when viewed from above. Deltoid ridge moderately prominent, irregular, rugose, and extending to about the middle of the shaft. Distal end of humerus not expanded. Bicipital groove inconspicuous.
Radius: Almost perfectly straight, but a little inclined toward ulna at oblique proximal end; scarcely expanded at distal end, which is lower externally than internally.
Ulna: Much slenderer than radius, rounded triangular in section, not expanded at distal end, where the margin is lowest externally. Olecranon well developed, thin, and pointed proximally.
Carpals: Five; two on ulna side, two median and one on radial side in line with first metacarpal. The proximal middle bone (intermedium) extends much farther proximally than those on each side of it.
Metacarpals: Metacarpal III longest, metacarpal II broadest. Metacarpal I oblong, or rather conical, with a lateral enlargement, and situated in line with the distal row of carpals.
Digits: First phalange of first digit short and conical.
Newport (male).—Fore limb considerably longer and more massive than that of the Barnegat skeleton but similar otherwise, except as follows:
Humerus: Head rather larger and less inclined. Deltoid ridge more prominent.
Radius: Broader proximally and rounded at distal end, where it extends outward beyond the carpal bones.
Ulna: Thicker, and olecranon less pointed.
Carpal bones: Middle carpal bone not extending farther proximally than those on either side of it.
Metacarpals: Metacarpal I nearly square, third longest, second to fourth more constricted.
Digits: First phalange of first digit long and cylindrical. Phalangeal formula: I, 1; II, 6; III, 6; IV, 4; V, 2.
Measurements of the skeletons above described are as follows:
Dimensions of four skeletons of Ziphius cavirostris.
| Measurements. | A | B | C | D |
|---|---|---|---|---|
| mm. | mm. | mm. | mm. | |
| Length of skull | 945 | 915 | 797 | ... |
| First to fourth cervicals (vert. 1-4): | ||||
| Length of combined centra | 81 | 79 | 66 | 55 |
| Greatest breadth of atlas | 283 | 259 | 210(?) | 146 |
| Greatest height of atlas | 215 | 218 | 170 | 148 |
| First thoracic vertebra (vert. 8): | ||||
| Greatest breadth | 211 | 220 | 174(?) | 122 |
| Greatest heighta | 267 | 321 | 163 | 133 |
| Length of centrum | 42 | 46 | 32 | 19 |
| Height of centrum | 70 | 67 | 48 | 49 |
| Seventh thoracic vertebra (vert. 14): | ||||
| Greatest breadth | 142 | 158 | 147 | 128 |
| Greatest height | 440 | 417 | 260 | 182 |
| Length of centrum | 102 | 103 | 74 | 51 |
| Height of centrum (ant.) | 68 | 69 | 49 | 44 |
| Eighth thoracic vertebra (vert. 15): | ||||
| Greatest breadth | 253 | 288 | 177 | 100 |
| Greatest height | 447 | 427 | 265 | 186 |
| Length of centrum | 110 | 109 | 78 | 54 |
| Height of centrum | 65 | 69 | 51 | 47 |
| Ninth thoracic vertebra (vert. 16): | ||||
| Greatest breadth | 329 | 366 | 248(?) | 122 |
| Greatest height | 418 | 431 | 277 | 192 |
| Length of centrum | 117 | 117 | 82 | 58 |
| Height of centrum | 69 | 75 | 53 | 49 |
| First lumbar vertebra (vert. 17): | ||||
| Greatest breadth | 385 | 393 | b275 | 142 |
| Greatest height | 464 | 451 | b293 | 200 |
| Length of centrum | 122 | 120 | b89 | 62 |
| Height of centrum | 74 | 81 | b55 | 50 |
| Tenth lumbar vertebra (vert. 26): | ||||
| Greatest breadth | 362 | 335 | c230 | 158 |
| Greatest height | 524 | 488 | c343 | 242 |
| Length of centrum | 172 | 162 | c129 | 85 |
| Height of centrum | 107 | 109 | c78 | 71 |
| First caudal vertebra (vert. 27): | ||||
| Greatest breadth | 336(?) | d313 | 223 | 166 |
| Greatest height | 483 | d458 | 307 | 235 |
| Length of centrum | 178 | d160 | 127 | 91 |
| Height of centrum | 113 | d109 | 81 | 75 |
| Seventh caudal vertebra (vert. 33): | ||||
| Greatest breadth | 164 | e161 | 113 | 110 |
| Greatest height | f255 | f250 | 155 | f135 |
| Length of centrum | 139 | 131 | 103 | 77 |
| Height of centrum (ant.) | f111 | f119 | f85 | f78 |
| Twelfth caudal vertebra (vert. 38): | ||||
| Greatest breadth | 90 | g88 | 61 | 62 |
| Greatest height | 83 | g78 | 53 | 61 |
| Length of centrum | 55 | g56 | 46 | 47 |
| Fifteenth caudal vertebra (vert. 41): | ||||
| Greatest breadth | 64 | h62 | 49 | 45 |
| Greatest height | 54 | h52 | 33 | 30 |
| Length of centrum | 42 | h39 | 30 | 28 |
| Eighteenth caudal vertebra (vert. 45): | ||||
| Greatest breadth | 37 | i38 | ... | ... |
| Greatest height | 20 | i19 | ... | ... |
| Length of centrum | 24 | i22 | ... | ... |
| Twentieth caudal vertebra (vert. 46): | ||||
| Greatest breadth | ... | j25 | ... | ... |
| Greatest height | ... | j13 | ... | ... |
| Length of centrum | ... | j19 | ... | ... |
| Chevrons: | ||||
| Antero-posterior length of first chevron | ... | 74 | ... | ... |
| Depth of same | ... | 66 | 44 | ... |
| Length of second chevron | 83 | 107 | 55 | 38 |
| Depth of same | 87 | 85 | 74 | 63 |
| Length of third chevron | 114 | 91 | 61 | 46 |
| Depth of same | 135 | 206 | 123 | 90 |
| Length of fourth chevron | 125 | 122 | 67 | 51 |
| Depth of same | 213 | 206 | 111 | 67 |
| Length of eighth chevron | 80 | 86 | 43 | 35 |
| Depth of same | 108 | 115 | 41 | 33 |
| Length of ninth chevron | 84 | 74 | ... | 27 |
| Depth of same | 83 | 88 | ... | 19 |
| Scapula: | ||||
| Greatest length | 385 | 415 | k224 | 159 |
| Greatest height | 275 | 300 | 175 | 132 |
| Length of acromion | l115 | 159 | ... | 48 |
| Length of coracoid from edge of glenoid | 127 | 148 | 60 | 38 |
| Pectoral limb: | ||||
| Total length | 588 | 652 | ... | ... |
| Humerus: | ||||
| Length | 168 | 177 | 130 | 95 |
| Breadth at distal end | 69 | 69 | 52 | 42 |
| Radius: | ||||
| Length | m178 | m175 | m135 | ... |
| Breadth at distal end | 55 | 65 | 41 | ... |
| Ulna: | ||||
| Length without olecranon | 165 | 171 | 118 | ... |
| Length including olecranon | 220 | 225 | 150 | ... |
| Breadth at distal end | 44 | 42 | 30 | ... |
| Metacarpals: | ||||
| Length of first | 31 | 28 | ||
| Length of second | 52 | 55 | ||
| Length of third | 52 | 58 | ||
| Length of fourth | 44 | 51 | ||
| Length of fifth | 37 | 39 | ||
| Phalanges: | ||||
| Length of first phalange of first digit | 27 | 44 | ||
| Sternum: | ||||
| Total length | n803 | 821 | o550 | o395 |
| Length of manubrium | 259 | 306 | o203 | 105 |
| Breadth of manubrium | 286 | 333 | 193 | 128 |
| Length of fifth segment | o170 | p184 | o128 | p92 |
| Breadth of fifth segment | 133 | 168 | 86 | 82 |
| Ribs: | ||||
| Length of first rib (straight) | 405 | 410 | 277 | 191 |
| Breadth of first rib at proximal end | 88 | 110 | 65 | 46 |
| Breadth of first rib at distal end | 63 | 80 | 40 | 30 |
| Length of fifth rib (straight) | 785 | 770 | 545 | 415 |
| Length of ninth rib (straight) | 620 | 620 | ||
The formulas for the ossified phalanges in two American[39] and three Old World specimens are as follows:
Phalangeal formula of five specimens of Ziphius cavirostris.
| Locality. | I. | II. | III. | IV. | V. |
|---|---|---|---|---|---|
| Newport, Rhode Island | 1 | 6 | 6 | 4 | 2 |
| Barnegat City, New Jersey | 1 | 6 | 6 | 4 | 3 |
| Villefranche, France (Haeckel) | 1 | 5 | 6 | 4 | 2 |
| Pisa Museum, Italy (Van Beneden) | 1 | 3(?) | 5 | 4 | 1 |
| Warrington, New Zealand (Scott and Parker) | 1 | 5 | 5 | 4 | 2 |
The chief differences between the Barnegat City and Newport skeletons are in the size and form of the processes of the cervical vertebræ, the form of the seventh and eighth thoracic vertebræ and of the ribs connected with them, the direction of the acromion of the scapula, the shape of the first phalange of the first digit, and of the posterior segments of the sternum. As far as the processes of the cervicals are concerned, these are known to be extremely variable in all cetaceans. The [50] seventh and eighth thoracic vertebræ are those on which the mode of attachment of the ribs changes in ziphioid whales, and I have observed in the genus Mesoplodon, as here, that the processes and articular facets were very variable, being sometimes quite unlike on the two sides of the same vertebra. The direction of the acromion is probably subject to large individual variations, though this can not be determined at present, and the same is true of the form of the first phalange of the first digit. The form of the sternum is quite variable in all cetaceans, and can not be relied on for specific characters, without comparison of many individuals.
On the whole, I am of the opinion, as already stated, that we are not compelled by the differences noted to regard the Barnegat and Newport skeletons as representing different species. The Charleston skeleton is too young and imperfect to admit of serious consideration. The idea that the differences between the adult skeletons are probably individual receives support from the fact that the skeleton shown in the photograph from St. Simon Island, Georgia, mentioned on page 31, No. 14, appears to possess a combination of characters exhibited by the other two.
The series of skulls of Z. grebnitzkii, which the Museum owes to the activities of Dr. L. Stejneger and Mr. N. Grebnitzki, comprises specimens of different ages, and, as will be shown presently, probably both sexes. Taken together with the skulls from the east coast of the United States they probably represent very fully the variations which the skull undergoes in the present species. These changes may, perhaps, be best made evident by the following brief descriptions of the various skulls:
21975. Charleston, South Carolina.—Young female. (Type of Z. semijunctus.) All sutures open, and elements of occipital bone distinguishable. No mesethmoid ossification. Opposite maxillary notches, premaxillæ closely approximated, nearly flat and horizontal, and about level with adjacent parts of maxillæ. Left premaxilla grooved longitudinally at this point. Orifice of anterior nares on a level with lower end of rectangular projecting boss formed by superior portion of nasals. Rostrum pointed, much broader distally than it is deep. A very distinct rudimentary alveolar groove in distal end of each maxilla. Proximal end of vomer resting against anterior face of nasals and reaching up to overhanging boss. Anterior face of the latter nearly flat. (Pl. 14, fig. 1; pl. 18, fig. 1; pl. 20, fig. 1; pl. 21, fig. 2.)
Rami of mandible not anchylosed together at symphysis. Teeth hollow, open at the root, acute at apex, tipped with enamel; diameter 10 mm. (Pl. 22, fig. 1; pl. 24, fig. 1.)
20971. Barnegat City, New Jersey.—Adult female. Majority of sutures open, but those on superior surface of rostrum between maxillæ and premaxillæ partly anchylosed. Vomer nearly all anchylosed to rostral portion of premaxillæ; it presents a slight median elevation, but there is no mesirostral ossification. Right premaxilla in front of nares broad, flat, and horizontal; left, nearly so, but with a quite broad longitudinal groove. Opposite maxillary notches premaxillæ nearly on a level with adjacent parts. Orifice of anterior nares level with lower end of nasal boss. End of rostrum quite acute, and broader than deep. Rudimentary [51] alveolar groove distinct distally. Proximal end of vomer anchylosed with anterior face of nasals and reaching up to nasal boss, which has a sharp median ridge completing nasal septum superiorly. Anterior face of nasal boss slightly concave on each side of median line. (Pl. 14, fig. 2; pl. 18, fig. 2; pl. 20, fig. 2; pl. 21, fig. 3.)
Rami of mandible anchylosed together at symphysis and suture largely obliterated. Teeth slender, cylindrical, rugose, rather blunt; roots closed; diameter 13 mm. (Pl. 24, fig. 3.)
22069. Bering Island.—Adult female? All the sutures about as in preceding specimen. Mesirostral ossification distinct, rounded, extending from base of rostrum nearly to apex, but disappearing before reaching line of anterior ends of maxillæ. Its upper surface below that of premaxillæ. Premaxillæ approximated, and right premaxilla with an angular process near base of rostrum overlapping mesirostral ossification. Premaxillæ at base of rostrum, anterior nares, proximal end of vomer, and nasals as in preceding skull. Apex of rostrum moderately acute, broader than deep. Rudimentary alveolar groove shallow. (Pl. 15, fig. 1.)
Rami of mandible anchylosed together and suture largely obliterated. Teeth somewhat fusiform, blunt; roots closed; diameter, 14 mm. (Pl. 22, fig. 3.)
83991. Bering Island.—Similar in all respects to preceding, but mesirostral ossification a little less well developed.
22874. Bering Island.—Entirely similar to two preceding, but premaxillæ a little curved out from mesirostral ossification and left premaxilla opposite maxillary notch rather strongly inclined, nearly vertical. Anterior face of nasal boss distinctly concave. (Skull defective.)
21246. Bering Island.—Sutures as in three preceding skulls. Mesirostral ossification distinct and rounded, but much below level of premaxillæ. Rostral portion of premaxillæ narrow and widely divergent toward base of rostrum, leaving mesirostral entirely exposed. Right premaxilla on a line with maxillary notches strongly concave and sunk below level of maxillæ. Left premaxilla vertical, with a broad groove. Right premaxilla remains low and concave proximally, the posterior end being then abruptly turned upward and reaching level of vertex. Orifice of anterior nares on a level with lower end of nasal boss, and vomer resting against anterior face of nasals, which latter have a median ridge continuing nasal septum, but with a slight vacuity between the two. Rudimentary alveolar groove nearly obliterated. Outer sides of premaxillæ at distal end strongly concave. Rostrum rather acute, about as deep as wide opposite distal ends of maxillæ. (Pl. 15, fig. 2.)
20993. Bering Island.—Adult male? (Type of Z. grebnitzkii). Majority of sutures open, but maxillæ and premaxillæ anchylosed together above and on the sides. Premaxillæ approximated anteriorly, but diverging posteriorly. Mesirostral ossification well developed, reaching level of premaxillæ; anteriorly rather narrow but a little broader near middle of rostrum, where it is beveled off abruptly. Behind this point premaxillæ strongly concave, nearly vertical and widely separated, forming a large and deep basin, in the bottom of which the vomer appears as a broad, irregular bony surface. Bottom of basin much below level of surrounding parts. Orifice of anterior nares much below level of nasal boss. Vomer reaching lower end of nasals. Anterior face of latter strongly concave, with only a moderate median [52] ridge completing nasal septum above. Mesirostral with a median groove at distal end. Premaxillæ high at distal end, but sides nearly plane. Rostrum compressed near apex, deeper than wide. (Pl. 16, fig. 1; pl. 19, fig. 1; pl. 20, fig. 3.)
Rami of mandible anchylosed together and suture partly obliterated. Teeth conical, with rather short, acute tips; roots closed, short and conical; diameter, 25 mm. (Pl. 23, fig. 1; pl. 24, fig. 2.)
21245. Bering Island.—Nearly all sutures between maxillæ and premaxillæ at end of rostrum, above and below, anchylosed together, but majority of others traceable. Condition of superior surface of skull very similar to that of preceding, but premaxillæ rather low at distal end. Mesirostral at distal end rather lower than premaxillæ and concave superiorly; more posteriorly assuming form of a narrow ridge, with a deep channel between it and premaxillæ on each side. More posteriorly still it widens rapidly, with a convex surface, and terminates abruptly with a truncated end, the surface of which is concave. A deep basin around nares, as in preceding skull. Orifice of anterior nares far below level of nasal boss. The latter largely absorbed and deeply undercut and concave in front. Nasal septum terminating before reaching lower end of nasals, and ridge on latter low and traversing left nasal. Sides of premaxillæ at distal end very concave. Rudimentary alveolar groove nearly obsolete. Rostrum blunt at apex, and about as deep as wide at anterior ends of maxillæ. (Pl. 16, fig. 2.)
21248. Bering Island.—Similar to preceding, but mesirostral ossification higher than premaxillæ at distal end and convex above; less abruptly widened posteriorly and posterior termination flat. Narrow, deep grooves between ossification and premaxillæ on each side, or, in other words, premaxillæ more closely approximated to sides of mesirostral distally. Basin around nares and conformation of the several bones bordering it similar to preceding. Sides of premaxillæ concave at distal end, the grooves thus formed in them intruding some what on the maxillæ, especially posteriorly. Apex of rostrum very blunt, rounded off below and projecting above; deeper than wide. Rudimentary alveolar groove nearly obsolete. (Pl. 17, fig. 1; pl. 22, fig. 4.)
Rami of mandible anchylosed together and the symphysis and suture largely obliterated. Teeth very broadly fusiform; tip short and rather blunt; roots closed; diameter 30 mm.
49599. Newport, Rhode Island.—Adult male. All sutures on superior surface of skull more or less anchylosed together. Mesirostral ossification and premaxillæ all on one level near apex of rostrum, but at extreme tip mesirostral lower, forming a narrow ridge with a deep groove on each side between it and premaxillæ. The same conformation repeated more posteriorly, but grooves deeper and wider, while mesirostral maintains the same level as premaxillæ. It widens suddenly here, forming a broad flat-topped mass, which is a little overlapped by the premaxillæ. The mass terminates suddenly somewhat behind middle of rostrum with a deep concavity placed obliquely. Basin in front of the nares and conformation of bones composing it as in two preceding skulls. Vomer at proximal end touching lower end of nasals, and nasal septum continued behind and above it as a low ridge, composed of the inner edges of the two nasal bones and reaching up to the nasal boss. Outer sides of premaxillæ near distal end deeply concave. Apex of [53] rostrum rather blunt, deeper than wide opposite distal ends of maxillæ; all the bones anchylosed together, but some of the sutures indicated by grooves. Rudimentary alveolar groove nearly obsolete. (Pl. 17, fig. 2; pl. 19, fig. 2; pl. 21, figs. 1, 5.)
Rami of mandible anchylosed together at symphysis, the suture indicated only by a groove. Teeth large, broadly conical and tapering at the tip. Root very short, rugose, conical and closed; diameter 29 mm. (Pl. 22, fig. 2; pl. 23, figs. 2, 3.)
The dimensions of the several skulls are as follows:
Dimensions of ten skulls of Ziphius cavirostris (including the types of Z. grebnitzkii Stejneger and Z. semijunctus Cope).
| Measurements. | A | B | C | D | E | F | G | H | I | J |
|---|---|---|---|---|---|---|---|---|---|---|
| mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | |
| Total length | 900 | 877 | a807 | 850 | 963 | 882 | 855 | 797 | 945 | 915 |
| Length of rostrum | 491 | 480 | a397 | 470 | 550 | 480 | 476 | 463 | 550 | 514 |
| Height from vertex to inferior border of pterygoids | 433 | 450 | ... | ... | 515 | 471 | 481 | 349 | 440 | 465 |
| Distance from tip of rostrum to posterior free margin of pterygoids (median) | 664 | 670 | ... | ... | 735 | 682 | 673 | 614 | 735 | 726 |
| Distance from the same to anterior end of nasals | 617 | 621 | a538 | 600 | 690 | 623 | 589 | 590 | 708 | 676 |
| Breadth between centers of orbits | 495 | 513 | b499 | 488 | 563 | b486 | 492 | 393 | 476 | 530 |
| Breadth between zygomatic processes | 511 | 513 | ... | 505 | 573 | 531 | 530 | 415 | 503 | 548 |
| Breadth between temporal fossæ | 270 | 309 | 325 | 300 | 349 | 317 | 311 | 242 | 302 | 313 |
| Breadth of rostrum at base | 319 | 331 | 345 | 324 | 380 | 337 | 320 | 249 | 307 | 337 |
| Breadth of rostrum at middle | 102 | 117 | b94± | 107 | 120 | 109 | 112 | 83 | 112 | 113 |
| Breadth of premaxillæ at same point | 54 | 67 | 58 | 62 | 78 | 70 | 75 | 44 | 62 | 80 |
| Depth of rostrum at middle | 66 | 81 | 80 | 79 | 118 | 117 | 113 | 50 | 77 | 107 |
| Breadth of premaxillæ in front of nares | 176 | 177 | 184 | 205 | 221 | 230 | 219 | 128 | 176 | 234 |
| Greatest breadth of anterior naresc | 74 | 77 | 77 | 90 | 98 | 103 | 108 | 70 | 76 | 112 |
| Greatest length of temporal fossa | 161 | 158 | 154 | 149 | 152 | 140 | 146 | 133 | 143 | 155 |
| Greatest depth of temporal fossa | 81 | 73 | 77 | 79 | 87 | 74 | 89 | 67 | 80 | 76 |
| Length of orbit (ant.-post.) | 131 | 133 | 132 | 130 | 137 | 126 | 117 | 113 | 134 | 132 |
| Distance from anterior end of orbit to maxillary notch | 78 | 92 | 82 | 70 | 83 | 89 | 85 | 61 | 82 | 99 |
| Length of tympanic bulla | ... | ... | ... | ... | 53 | ... | ... | 54 | ... | 55 |
| Breadth of tympanic bulla | ... | ... | ... | ... | 24 | ... | ... | 37? | ... | 25 |
| Length of mandible | ... | 769 | ... | ... | ... | ... | ... | 679 | ... | 842 |
| Length of symphysis | ... | 170 | ... | ... | ... | 184 | ... | 149 | ... | 176 |
| Depth of mandible at coronoid | ... | 153 | ... | ... | ... | ... | ... | 133 | ... | 153 |
It will be found from an examination of the foregoing descriptions that in those specimens in which the sex is known to be female, or is marked as such, the premaxillæ are comparatively narrow, the mesirostral ossification only slightly developed, the prenarial basin undeveloped, and the teeth quite slender, with a diameter of from 10 to 14 mm. As the teeth in some of them have closed roots there can be no doubt that they are adults. On the other hand, those skulls known or believed to be from adult males have the mesirostral ossification enormously developed, a deep prenarial basin, and fusiform teeth with closed roots and a diameter of from 25 to 30 mm. It appears to be a fact, therefore, that in the females the mesirostral ossification is never greatly developed at any age, that the teeth are never thick and fusiform, and that the prenarial region is never deeply concave. Immature individuals present, of course, the appearance of the females, except that the teeth are open at the root and that the mesirostral ossification is not developed at all. Conversely, the females, broadly speaking, always present characters of immaturity, but in adults the roots of the teeth are, of course, closed.
That these conclusions are correct is borne out by an examination of descriptions and figures of specimens from other parts of the world, for which purpose a few are available in the writings of New Zealand zoologists and others. Hector, for example, in 1873,[40] published a description and figures of a skull from the Chatham Islands which had a large mesirostral ossification, deep prenarial concavity, and large, thick teeth, having a diameter of 34 mm. This is the same combination of characters found in the Newport specimen, which is known to be a male, and the Bering Island skulls supposed to be those of males.[41]
In 1876,[42] Haast figured and described a female 26 feet long, and hence presumably adult, from Lyttleton Harbor, New Zealand, which had a small development only of the mesirostral ossification, a slight prenarial depression, and rather slender teeth with closed roots and a diameter of 19 mm. This combination of characters is found in the Barnegat skull, also known to be an adult female.
In the same paper Haast describes[43] and figures the skull of another female from Akaroa Harbor, New Zealand. This individual was larger than the last and was accompanied by a suckling calf. Hence, there can be no doubt that it was mature. The skull shows a moderate development of the mesirostral ossification, and slender cylindrical teeth with closed roots and a diameter of 16 mm.
It is demonstrated from the foregoing discussion, I think, that the sexes can be distinguished by the skulls, when adult, or by the teeth alone.
Reverting now to Ziphius gervaisii, which was mentioned on p. 30 as perhaps constituting a separate species, it will be seen by examining the figures given by [55] Gervais[44] of the skull on which it was based that the latter presents the combination of characters peculiar to the female of Z. cavirostris. This skull, which was from Aresquiers (Hérault), France, was 888 mm. long, and hence, presumably, adult. The mesirostral ossification is but slightly developed, the prenarial concavity moderate, the teeth small, slender, and cylindrical, with closed roots and a diameter of 14 mm. There seems to be no sufficient reason for regarding this skull as representing a species distinct from cavirostris.
The specimen from Buenos Ayres described and figured by Burmeister in 1868[45] was an immature male. In the skull the mesirostral ossification was lacking, the premaxillæ were flat, and the teeth conical and acuminate, with open roots, and a diameter of 12 mm. This individual was 12 feet 11½ inches (3.95 m.) long, and hence about as long as the Charleston specimen, but the skull was apparently 680 mm. long, while that of the Charleston specimen is 797 mm. long. In the latter the teeth are 45 mm. long and 10 mm. in diameter, while the tooth figured by Burmeister is 31 mm. long and 12 mm. in diameter. From these data it appears improbable that the sex of immature individuals can be determined from the skull or teeth.
The teeth of the various North Atlantic and North Pacific specimens merit a somewhat more detailed description than is given on pages 50 to 53. Six pairs of teeth from six different individuals are available for comparison. Their dimensions are as follows:
Dimensions of the teeth of Ziphius cavirostris.
| Cat. No. | Locality. | Age. | Sex. | Teeth. | |
|---|---|---|---|---|---|
| Length. | Greatest diameter. | ||||
| mm. | mm. | ||||
| 21975 | Charleston, South Carolinaa | Young | Female | 45 | 10 |
| 20971 | Barnegat City, New Jersey | Adult | Female | 56 | 13 |
| 22069 | Bering Island | Adult | (Female?) | 41 | 14 |
| 20993 | dob | Adult | (Male?) | 48 | 25 |
| 21248 | do | Adult | (Male?) | 58 | 30 |
| 49599 | Newport, Rhode Island | Adult | Male | 63 | 29 |
21975. Charleston, South Carolina.—Young female. (Type of Z. semijunctus.) The teeth are slender, conical, and acuminate, largest at the base and tipped for about 2 mm. with white enamel. The remainder of the teeth is coated with a thin layer of cement. The teeth in what appears to be their natural position protrude horizontally from the mandible for about 17 mm. They are slightly curved upward near the tip and are oval, or elliptical, in section, the transverse diameter being a little less than the vertical diameter. They are a little flattened externally. [56] The surface is smooth. They are open at the root, and hollow. (Pl. 38, figs. 1, 2; pl. 22, fig. 1.)
Doctor Manigault, curator of the Charleston Museum, wrote to Professor Cope regarding these teeth, as follows:
Another peculiarity of the head consists in the lower maxillary bones being provided each at its point with a single small and very sharp tooth. These were not noticed during the dissection, owing to their being too much embedded in the integuments.[46]
20971. Barnegat City, New Jersey.—Adult female. The teeth are slender, cylindrical, and irregularly pointed at both ends. The tips show what appears to be an inner core of dentine which has been worn down nearly to the cement coating and somewhat fractured. The cement coating is several millimeters thick, but does not increase the diameter of the teeth near the middle, so that they remain irregularly cylindrical throughout. The surface of the cement is rough and irregular. The root is short, conical, and closed at the end. These teeth are nearly straight. As they have been extracted from the jaw and the latter is broken it is not possible to distinguish which is the upper and which the lower surface, but they are irregularly oval in section, and a little compressed. (Pl. 38, figs. 3-5.)
In my original notes on this specimen, I recorded that there was a small pair of teeth behind the larger ones described above. Mention of these will be made again later. (See p. 57.)
22069. Bering Island.—Adult female (?). The teeth are in position in this specimen and are nearly horizontal in position, but a little inclined upward and toward each other. They do not extend beyond the tip of the jaw nor up to the level of the upper surface of the symphysis, but protrude about 13 mm. beyond the alveoli on the side. They are rather slender, somewhat fusiform, blunt at both ends and slightly curved upward. The surface is irregular. They are nearly round in section. The root is closed, and the apex shows what appears to be a core of dentine surrounded by cement. There is a depression on the inner side near the root. These teeth are remarkable as intermediate in form between those of the preceding specimen and those of the specimens next to be mentioned. (Pl. 38, figs. 6, 7; pl. 22, fig. 3.)
20993. Bering Island.—Adult male (?). (Type of Z. grebnitzkii.) These teeth are almond-shaped and very symmetrical. They are thickest near the base and taper gradually to the tip, which is quite acute. They are somewhat compressed and hence elliptical in section, the vertical diameter being greater than the transverse diameter. One side (probably the inner) is flattened. They are slightly curved upward toward the apex, which is a little worn and fractured. The root is very short and conical. It is nearly closed, but a very small canal extends upward for about 10 mm. The surface of the tooth is quite smooth, but dull in the lower half. The line of demarcation between cement and dentine is not evident. (Pl. 38, figs. 8, 9; pl. 23, fig. 1.)
21248. Bering Island.—Adult male (?). In this specimen the teeth are still in the natural position in the jaw. They are held in place by ligaments and protrude [57] far beyond the alveoli, only about one-ninth of their length being below the superior border. They incline forward at an angle of about 45° with the longitudinal axis of the jaw and diverge slightly at the tips.
The teeth themselves have the same general form as those of the preceding specimen, but are larger. The inner surface is flattened and the outer strongly convex. The tips are quite pointed, but show some indications of wear. The roots can not be seen distinctly, but appear to be closed. (Pl. 22, fig. 4.)
49599. Newport, Rhode Island.—Adult male. These teeth are longer than those of the preceding specimen, and while they resemble the latter in general form, taper much more gradually to the tip. The root, or portion below the point of maximum girth, is much shorter than that above, and rugose, with several deep furrows. A very small circular opening at the base of the root marks the orifice of the nerve. The upper half of the teeth is smooth, and the tips slightly worn and fractured. The small elliptical worn area is situated on the convex side of the tooth, which appears to be the outer side. As the alveoli of the jaw are, however, filled with a network of bone, the teeth can not be inserted in them. They were detached when received. (Pl. 38, figs. 10, 11; pl. 22, fig. 2; pl. 23, figs. 2, 3.)
Besides the difference in the size and form of the teeth in the two sexes, it is probable, as will be seen by consulting the foregoing data, that in the female the apex of the teeth does not extend more than a very small distance above the alveoli even in mature individuals, and probably often not more than a few millimeters; while in adult males the teeth are almost entirely protruded from the alveoli, which are filled with a coarse bony network. These differences are carried out in all the American specimens, and also characterized the New Zealand specimens, as may be learned from the accounts of Haast and Hector.
A number of rudimentary teeth in addition to the large terminal pair have been noted in the Aresquiers, Buenos Ayres, and perhaps other specimens, and two such teeth were found in the mandible of the Barnegat specimen, behind the large pair. One of these rudimentary teeth has been preserved. It is cylindrical and moderately curved. The length is 16 mm. and the diameter 2 mm. The whole tooth, with the exception of the extreme tip, is thickly coated with cement. The root is closed and the crown acute and apparently abraded by use. (Pl. 38, fig. 5.)
Returning now to the question of the validity of grebnitzkii as a species, I would say that after comparing the measurements of the Bering Island skulls with those of the Atlantic coast specimens, and comparing the skulls themselves, I have been unable to find any constant difference of importance, except the size and form of the periotic bone. As the earbones are lacking from many of the skulls, the series available for comparison is small.
As compared with the Atlantic coast specimens, the anterior portion of the periotic bone in grebnitzkii is larger, broader, and more rectangular in outline when viewed from below. I observe, however, that the absolute size and outline of the periotic vary considerably in the different specimens of grebnitzkii without relation to age. The same appears to be true of cavirostris, but comparing the two series of skulls as a whole it appears to be true that the anterior mass of the periotic is larger in grebnitzkii. I do not think, however, that the latter species should be [58] kept distinct on this account alone, at least until the character has been confirmed, and perhaps strengthened by others, through the examination of a larger series of specimens.
The Museum collection contains an incomplete skeleton of a very young individual, Cat. No. 22875, which was received from Bering Island with the skulls of Z. grebnitzkii, but does not belong to any one of them. Whether it really represents that species is, therefore, uncertain, but such is probably the case. The length of the vertebral column, consisting of 45 vertebræ, without interspaces, is 9 feet 2 inches.
The vertebral formula is as follows: C. 7; Th. 10; L. 10; Ca. 18 (+1?) = 45 (+1?). This is the same as in the type of semijunctus so far as the cervicals, thoracics, and lumbars are concerned, and the probable total is the same. In their general characters these vertebræ agree with those of the skeletons already described, but they present a number of differences as well. On account of immaturity the processes are even less developed than in semijunctus. All the epiphyses are free, and in the third to the seventh thoracic vertebræ the neural arch and spine are separate from the centrum. The centra are very short in proportion to their width.
Although the specimen is so young, the anterior foramen of the atlas is, nevertheless, inclosed by bone, and though the line of separation between the atlas and axis is visible on the sides, the fourth cervical is anchylosed to the third at the top of the centrum. Although the neural spines, metapophyses, and transverse processes of the thoracics are much shorter than those of the young semijunctus, the epiphyses are as large or even larger than in that specimen. The neural arches are also noticeably thicker than in semijunctus, and the centra are rounded inferiorly rather than carinated. The neural spines are much more nearly erect than in the adult Barnegat and Newport skeletons, but, as mentioned on page 41, this is probably a character of immaturity, and is shared by semijunctus.
The differences as regards the form of the centra and neural arches die away among the lumbars, and these vertebræ and the caudals are, with a due allowance for greater immaturity, very similar to those of semijunctus.
The seventh thoracic is like the sixth in form, and is without a transverse process. It thus resembles the same vertebra in semijunctus. The eighth, however, has an ill-defined facet on the side of the metapophysis and a second facet a little above the upper border of the centrum. The eighth pair of ribs has only a single terminal articular facet.
The ninth thoracic has a short, thick transverse process, about in line with the upper surface of the centrum.
The transverse process of the seventh caudal is perforated on the right side by a foramen. The transverse processes are last traceable on the ninth caudal, the neural spines on the tenth caudal, and the neural arch on the eleventh caudal. Eight chevron bones are preserved, but probably two more were present originally.
Ten pairs of ribs are present. The first is much broader in the proximal half than in the distal half, but the distal end is slightly expanded. The first seven pairs [59] possess both head and tubercle, but the eighth, ninth, and tenth have only a single terminal articular facet.
The sternum, which consists of five segments, is similar in form to that of semijunctus. The two sides of each segment are united. The posterior emargination of the third segment, and those of both ends of the fourth and fifth segments are small. The scapula and humerus are like those of semijunctus in form. The remaining parts of both pectoral limbs are lacking.
Without more material, and especially some skeletons of adults, it is difficult to decide what importance should be assigned to the differences observable in the cervical and thoracic vertebræ of this young Bering Island specimen. The measurements of the skeleton are included in the table on pages 47 and 48.
The series of photographs (Cat. No. 142579) of an individual obtained in Kiska Harbor, Alaska, is very interesting as affording comparison of what is apparently a specimen of grebnitzkii with the Atlantic form represented in the photograph of the Newport, Rhode Island, specimen. As no part of the Kiska specimen was preserved, it is not possible, of course, to identify it positively with grebnitzkii or even with the genus Ziphius. No one who compares the photographs reproduced in Pl. 41, figs. 3 and 4, can, I think, fail to be convinced that both represent animals of the same genus and that the Pacific species (whether grebnitzkii or not) bears the strongest possible resemblance to the Atlantic one.
Doctor Egbert published the following note on the Kiska specimen in 1905:
Early in September a monster dolphin grounded on the beach in Kiska Harbor and was killed. Specific identification has not yet been made. The general color was bluish-gray; length, 18½ feet; estimated weight, 3,600 pounds; sex, male. Body was quite regular in shape and rather rotund, the greatest circumference being about midway between dorsal fin and tip of the rather short snout. This dolphin was hauled alongside the ship, stripped of its blubber, and the oil extracted. Some of the flesh was eaten. The oil obtained was of excellent quality. It was particularly desired for use on the wire of the deep-sea sounding machine used aboard the [U. S. Coast Survey steamer] Patterson.[47]
The size was about the same as that of the Newport specimen. Although Doctor Egbert gives the color merely as “bluish gray,” the photographs indicate that the belly was white, or whitish, and that there were oval white spots on the sides. As a whole, therefore, the coloration was similar to that of the New Zealand specimens of cavirostris obtained at Port Cooper and Lyttleton Harbor.
When compared with the photograph of the Newport specimen (Pl. 41, fig. 4) it will be seen that the Kiska photograph represents an animal practically identical in general form, as well as in the general shape of the head, the length and form of the snout, the size and general shape of the pectoral fins. In the photograph of the Newport specimen the flukes are not well seen, but in the Kiska photograph the posterior median convexity peculiar to the ziphioids is clearly represented. The dorsal fin of the Newport specimen appears to be turned somewhat to one side and the tip crumpled, which makes it appear lower and somewhat longer and less pointed than [60] that of the Kiska specimen. This may, of course, be a real difference, though such is probably not the case.
Considering the foregoing data relative to grebnitzkii as a whole, there is not in my opinion sufficient warrant at present for considering this form as a species distinct from cavirostris, and it should be added that no distinguishing characters were given in the original description.
Of this genus the National Museum has three skulls and three skeletons representing the species bairdii, and a skull representing the species arnuxii. The latter, Cat. No. 21511, U.S.N.M., is without exact locality, but is catalogued as having been obtained in New Zealand. As the species arnuxii has been well described and figured by Flower[48] and others, no detailed account of this skull is given here. Measurements of it, however, are included with those of B. bairdii in the table on p. 68.
This species was based by Dr. L. Stejneger on a skull obtained by Mr. N. Grebnitzki in Stare Gavan, on the eastern shore of Bering Island, Commander Group, Bering Sea, in the autumn of 1881. In 1879 a portion of a skull of the same species was found on Bering Island by the Vega expedition, and was made the basis of a new species, B. vegæ, by A. W. Malm, the description of which was published a few months after that of Doctor Stejneger. The National Museum subsequently received another skull from Bering Island, through Mr. N. Grebnitzki, but, so far as I am aware, nothing further was heard of the species until 1903 and 1904, when the National Museum received three nearly complete skeletons, two of them from St. George Island, Pribilof Group, Bering Sea, and one from the coast of California. The material now in the National Museum is as follows:[50]
(1) Cat. No. 20992.—Skull and mandible of an immature individual collected by Dr. L. Stejneger in Bering Island. Original number 1520. Catalogued November 24, 1883. Type.
(2) Cat. No. (lacking).—Skull and mandible of an immature individual. Collected by Mr. N. Grebnitzki in Bering Island (?). Mounted.
(3) Cat. No. 142118.—Skull, mandible, and cervical vertebræ of a very young individual. Collected by Dr. L. Stejneger, June 5, 1883, on North Rookery, Bering [61] Island. Original number 2191. This specimen is accompanied by notes and measurements.
(4) Cat. No. 49726.—Skeleton and measurements of an adult female. Near East Rookery, St. George Island, Pribilof Group. Collected by James Judge, in June, 1903. Length, 40 feet 2 inches.
(5) Cat. No. 49727.—Skeleton and measurements of an immature male. Same locality and date as the preceding. Length, 25 feet 5 inches.
The two skeletons (4) and (5) are somewhat incomplete. The Museum received a photograph of the female from Maj. Ezra W. Clark.
(6) Cat. No. 49725.—Skeleton and two photographs of an adult male (?) stranded on Centerville beach near Ferndale, Humboldt County, California, October, 1904. Length, about 41 feet.
A brief note on the St. George Island and California skeletons was published by the author in Science for 1904.[51] The dimensions given by the collectors were so large as to raise doubts whether they were correct, but the arrival of the skeletons proved that they were not overstated, and that the specimens were by far the largest ziphioid whales ever discovered, the bones about equaling those of a humpback whale in size and massiveness.
The St. George Island specimens were first made known by Mr. James Judge, special agent of the Treasury Department, resident at the Pribilof Islands, in a letter dated June 16, 1903, as follows:
I was much surprised the other day to find a pair of whales ashore near East Rookery [St. George Island]. They lay about 150 yards apart. The female was 40 feet 2 inches, the male 25 feet 5 inches in length. The species is not positively identified, but tallies closely with the Globe Encyclopedia description of Bottlehead or Bottlenose whale, Hyperodoön bidentatus. Natives call it “Tcha-dhan.” The male is without teeth; female has two teeth in front of lower jaw.[52] The skin is thin, smooth, white underneath, and black above. Dorsal fin small and well aft. Caudal large and powerful. Eyes very small. Ears not visible.
Thinking that the skeleton might be of use, the bones of the female were cut out and placed high and dry on the grass. Four ribs were broken; otherwise the bones are intact. The male was towed to East Landing, and with the aid of a capstan deposited beyond reach of surf. Some blubber was saved. The foxes will clean up the bones during August, so that in all probability both skeletons will be available this fall. * * * I inclose some measurements, taken roughly, with a 5-foot tape line.
Whale measurements, June 11, 1903.
| Female. | Male. | |||
|---|---|---|---|---|
| Ft. | in. | Ft. | in. | |
| Greatest length | 40 | 2 | 25 | 5 |
| Greatest circumference (much bloated) | 20 | 0 | 12 | 0 |
| Extremity of upper lip to nostril | 4 | 4 | 3 | 0 |
| Distance between eyes | 4 | 6 | 3 | 6 |
| Extremity of lower lip to angle of mouth | 2 | 5 | 1 | 9 |
| Circumference of head at eyes | 8 | 10 | 7 | 0 |
| Lower half of snout 10 inches from end | 2 | 3 | 1 | 9 |
| Upper half of snout 12 inches from end | 2 | 1 | 1 | 7 |
| Length of [pectoral] fin along outer edge | 5 | 0 | 3 | 5 |
| Circumference of tail [at] junction [with] caudal fin | 5 | 0 | 3 | 5 |
| Distance between extreme points of caudal fin | 10 | 2 | 6 | 3 |
| Anus to end of body | 11 | 8 | 7 | 7 |
| Anus to vagina | 1 | 2 | ... | ... |
| Anus to penis | ... | ... | 1 | 8 |
| Length of vagina | 1 | 3 | ... | ... |
| Length of penis | ... | ... | 1 | 9 |
| Penis at base | ... | ... | 1 | 5 |
| Height of dorsal fin | 0 | 12 | 0 | 7½ |
| Dorsal fin along spine to end of body | 11 | 11 | 7 | 5 |
| Length of nipple from raised base | 0 | 1 | ... | ... |
The skeletons remained on the island until August, 1904, when they were carried by the revenue cutter McCulloch to Dutch Harbor and afterwards to San Francisco. Through a misunderstanding they were allowed to remain on the beach at St. George Island until November, 1903, and suffered considerable injury. On that date they were deposited in a storehouse by Maj. Ezra W. Clark, assistant treasury agent in charge, who afterwards presented the photograph of the female above mentioned. (Pl. 42, fig. 1.) The latter shows the short, narrow, pointed pectoral fin, and long, rather slender beak.
Another specimen of Berardius was found stranded on St. George Island on August 21, 1909. The following information regarding it was received from Maj. Ezra W. Clark, under date of September 4, 1909:
On August 21, 1909, after an unusually severe gale for the season, accompanied with heavy sea, a beaked whale was stranded under the cliffs of the northeast coast of St. George Island. Its position was such that it was reached with great difficulty. It was undergoing decomposition. I succeeded in getting the following information:
I think that I shall not be able to get the skeleton of this whale, owing to the rough seas prevailing.
The Californian specimen (Cat. No. 49725) was first made known in a letter addressed to me by President Jordan, of Stanford University, under date of October 27, 1904, inclosing one from Mr. J. H. Ring, of Ferndale, California, dated October 23, 1904, which was as follows:
Enclosed find three views of an animal stranded on the beach near this place [Ferndale, Humboldt County, California], and as its identity seems rather uncertain we hope you will kindly classify it and inform us of its true name and habitat, if possible, from the photographs and incomplete description. Its total length is about 41 feet. Greatest circumference 16 feet, tapering probably to 18 inches near the tail. It also tapers toward the head, terminating in a sharp beak, the upper jaw being about 16 and the lower 19 inches long.
On each side in the lower jaw well to the front is a conical tooth, the crown of which is exposed one-half an inch. The head is full and rounded, resembling that of an elephant, with depressions corresponding to the ears, and small eyes a little ahead and below.
On top of head is a heart-shaped opening, evidently for breathing purposes. There is also evidence of a dorsal fin, while each fork of tail is 3½ feet or so long. The underside of the animal is too bruised to show anything of importance. The flippers are also in bad shape, one being buried in the sand, while the other is entirely denuded of flesh, leaving a bony stump about 6 inches long and which moves readily in any direction. We think it is a “bottle-nose” whale, but as some claim that they are not to be found on this coast and do not exceed 30 feet in length, it may be something else.
Mr. Ring was immediately communicated with, and very generously presented to the Museum the skull of the animal, which he had secured and cleaned with much labor and some danger to himself. He also undertook to have the skeleton cleaned and sent to Washington, and it was received in due course in June, 1905. Mr. Ring wrote under date of May 15, 1905:
You will notice that the point of the beak, as well as the points of the lower jawbones, are a little damaged, some hunters having shot the teeth out and then set a fire inside the jaws.
When received, the skeleton lacked the flippers and also two of the teeth. Regarding the former, Mr. Ring wrote on November 18, 1905, as follows:
I wrote you that one flipper was entirely gone and the other worn down to a stump, as shown in the picture. I have interviewed the man who stripped the specimen, and he says the stump was badly crushed and broken and fears it was lost one night when the extremely high tide had turned the whale over, and only the anchors and lashings I had secured it with prevented its going out to sea.
This skeleton was mounted recently and placed on exhibition in the Museum. The flippers were modeled from those of the St. George Island specimens (which were also imperfect) and from the figures of B. arnuxii given by Flower. The end of the beak was also restored, and a facsimile of the teeth substituted for the real ones. This remarkable skeleton shows in a manner hitherto unapproached the great size which this genus of ziphioid whales attains, and the peculiar conformation of the body. While the vertebræ rival those of the large whalebone whales, such as the Humpbacks, in their dimensions, the head is remarkable for its small size as compared with the immense proportions of the same part in the Right whales. (Pl. 42, fig. 4.)
Mr. Ring sent to the Museum three photographs of the Californian specimen above mentioned, two of which are reproduced on Pl. 42, figs. 2 and 3. Although rather indistinct, they show the general form of the body, the peculiar bulbous head, with an indication of a neck, and the long beak.
Doctor Stejneger has very kindly placed in my hands his original notes on the young individual examined by him in Bering Island June 5, 1883 (Cat. No. 142,188) and they are given below in full:
When the news reached me that a small “plavum” was found dead ashore at the North Rookery of Bering Island, I immediately ordered dogs, and arrived at the place in company with the “starost.” The carcass was found lying on the very beach where the fur seals during the summer occupy the ground. As the bulk of the seals had not yet arrived, only a few “sikatschi” were seen in the immediate neighborhood, but it was reported that they had retired from the place on account of the smell of the putrefied body, as it was thought. The natives, fearing that it would drive the seals from the rookery altogether if left on the beach any longer, were very anxious to get it away as fast as possible, and it was only with some hesitation that they would allow one to stand on the rookery long enough to take a few measurements. The animal was quite a young one, and I conjectured that it had died immediately after having been born, as I think there were some remains of the umbilical cord. Hardly any of the bones were fully ossified. Under these circumstances, it was out of the question to have the whole skeleton preserved, as the dismembering and the separation of the putrified flesh from the bones and cartilages would require more care and consequently more time than the natives were willing to allow. I was therefore glad to secure the head and some of the neck vertebræ. Even that tried their patience, as the head was going to separate into its single bones and the not yet united component pieces, and consequently needed special care and attention.
The carcass was lying with the back upward, this visible part being uniform black, and still in such a state as to allow of measuring. The lower surface was in a very advanced state of decomposition. Part of the belly was torn away, together with the entrails, and the genitalia and anus were not to be found. As stated above, I think that I could recognize the umbilical cord attached to a tatter of the skin. Of course, measurements of the lower side and of the circumference of the body, except at the narrowest place of the tail, could not be taken.
Table of dimensions.
| Meters. | |
|---|---|
| Total length from tip of upper jaw to notch of caudal fin, along the middle of the back, without, however, following the angle between beak and forehead | 4.81 |
| From tip of upper jaw to fore border of spiracles | .53 |
| From fore border of the spiracles to fore border of dorsal fin | 2.63 |
| Length of dorsal fin | .29 |
| Height of dorsal fin | .11 |
| From hind border of dorsal fin to the beginning of the caudal fin | .93 |
| From the same point to notch of the caudal fin | 1.36 |
| Distance between the tips of the lobes of the caudal fin | .91 |
| Depth of the angle of the posterior margin of caudal fin | .20 |
| From tip of upper jaw to the angle of mouth | .36 |
| From the same to anterior angle of eye | .475 |
| Diameter of eye opening | .06 |
| From eye to eye over the spiracle | .59 |
| Distance between ends of spiracle | .08 |
| Length of beak from the forehead | .23 |
| Breadth of the beak at the forehead | .18 |
| From tip of upper jaw to anterior insertion of the pectoral fin | .80 |
| Pectoral fin along the anterior border | .51 |
| Breadth of pectoral fina at the insertion | .20 |
| Circumference of tail at its narrowest point, just before the caudal fin | .62 |
The original description of B. bairdii by Doctor Stejneger is as follows:
Besides an Orca, which is said to visit the rookeries, but of which I have not been able to procure any specimen, or even to see one, there are at least two species of the family Ziphiidæ, both undescribed, as I suppose. I am very much indebted to Mr. Grebnitzki for a skull of each of the species, for one of which I should like to propose the name Berardius bairdii, as a slight token of my esteem and gratitude.
As I am now almost without any literary means, I find it impossible to decide with certainty in what genus this species will finally have to be placed. But I think that the supposition that this specimen (No. 1520) is a young Berardius may not be far out of the way. At first I suspected that it is a Dioplodon, but the size of the skull, in connection with the distinctness of the sutures, the evident maxillary crests, and the terminal position of the teeth very soon led me to the above conclusion.
The specimen in question has very low and scarcely incurved maxillary crests; the shortest distance of which is two and two-thirds times greater than their greatest height, and although it still is in its “adolescent” stage, I should greatly doubt whether the crests in this species ever become developed to such a degree as, for instance, in Hyperoödon diodon (Lacép.). The groove between the maxillary and the nuchal crest is very shallow. The maxillary notch is deep. The beak is long, making only a little less than half the length of the entire skull. Nares straight; right nasal larger than the left one, but not very much. The occipital condyles do not come in contact beneath the foramen magnum; the symphysis of the lower jaw is very short, amounting to only one-fifth of the whole length of the jaw.
Want of time and books prevents me from making more extended remarks, and until I can present an exhaustive and comparative description, I shall have to content myself by giving a provisional table of dimensions. The following dimensions are in millimeters and English inches, and are in every case measured in a straight line:
mm. in. Length of skull 1,405 55.32 Greatest breadth 698 27.48 Greatest height 530 20.87 Length from process of supramaxillaries before orbit to posterior edge of condyles 610 24.02 Length from same process to tip of beak 890 35.04 Depth of maxillary notch 50 1.97 Length of premaxillaries 1,222 48.11 Premaxillaries reach beyond supramaxillaries 134 5.28 Distance of upper edge of maxillary crests at their anterior end 228 8.98 Distance of same at their middle 358 14.10 Greatest height of maxillary crests 86 3.39 Length of visible part of vomer 325 12.80 Distance from anterior tip of vomer to tip of beak 275 10.83 Length of pterygoids 295 11.62 Height of foramen magnum 70 2.76 Width of foramen magnum 80 3.15 Distance of condyles at upper edge of foramen magnum 100 3.94 Closest approximation of condyles beneath the foramen magnum 2 0.08 Entire length of lower jaw 1,292 50.88 Height of lower jaw at second tooth groove 100 3.94 Length of symphysis 257 10.12 Greatest diameter of foremost tooth groove (longitudinal) 100 3.94 Shortest diameter of foremost tooth groove (transverse) 45 1.77 Greatest diameter of posterior tooth groove (longitudinal) 40 1.58 Shortest diameter of posterior tooth groove (transverse) 35 1.38 Distance between the tooth grooves 65 2.56 [66]This specimen was found stranded in Stare Gavan, on the eastern shore of Bering Island in the fall of last year, and only the skull was preserved. From analogy I should judge that the entire length of the animal must have been about 18 feet (5½ meters). This species is well known by the natives for the cathartic quality of the blubber, resembling in this respect the Atlantic “Dögling,” or “Anarnak” (Hyperoödon diodon). The Russian name, by which the inhabitants here designate this whale, is Pla-un (sp. Pläoon), while the Aleut name is Kigan agalusoch, the meaning of which is said to be “having teeth on the nose,” a very inappropriate designation, as the teeth are situated on the tip of the lower jaw, and not on the nose.[53]
It will be observed that the largest of the foregoing specimens measured 40 feet 2 inches in length, while the Centerville skeleton was reported to be about 41 feet long. The largest example of the New Zealand species, B. arnuxii, of which there is a record was 32 feet long.
The St. George Island specimens were reported to be black on the back and white below, but it is not certain how long they had been dead when found by Mr. Judge. The young individual examined by Doctor Stejneger was also black on the back, but this was in a state of decomposition.
The color of the type-specimen of Berardius arnuxii was described by Arnoux as follows: “Its color was entirely black, except for a light gray area near the genital organs; it was a male.”[54] Haast remarks of a young individual observed by him near New Brighton, New Zealand, and not in a fresh condition: “The color of the whole animal was of a deep, velvety black, with the exception of the lower portion of the belly, which had a grayish color.”[55]
The color of the immature male of B. arnuxii captured in Wellington Harbor, New Zealand, in 1877, and described by Hector, was as follows: “The colour was black with a purple hue, except a narrow band along the belly, which was grey. The muzzle, flippers, and tail lobes were intensely black.”[56]
It is not likely that there is any marked difference in the color of arnuxii and bairdii, but the data available are insufficient for the determination of the matter. It will be observed, however, that Mr. Judge stated that the male bairdii found on St. George Island was white below, while in all the accounts of arnuxii the color of the under surface is given as blackish, with a restricted area of gray.
Besides its apparently greater size, Berardius bairdii differs from B. arnuxii in various cranial and other osteological characters, as well as in external proportions, and is to be regarded as a distinct species. The external measurements of the St. George Island specimens reduced to percentages of the total length and compared with similar measurements of a specimen of B. arnuxii described by Hector, are as follows:
External dimensions of Berardius bairdii and B. arnuxii.
| Measurements. | A | B | C | |||
|---|---|---|---|---|---|---|
| ft. | in. | ft. | in. | ft. | in. | |
| Total length | 40 | 2 | 25 | 5 | 27 | 6 |
| per cent. | per cent. | per cent. | ||||
| Distance from tip of snout to blowhole | 10.8 | 11.8 | 12.8 | |||
| Distance from tip of mandible to corner of mouth | 6.0 | 6.9 | a6.1 | |||
| Breadth of flukes from tip to tip | 25.3 | 24.6 | 19.1 | |||
| Length of pectoral fin along outer edge | 12.4 | 13.4 | 9.4 | |||
| Distance from anus to “end of body” | 29.0 | 29.8 | [34.0] | |||
| Height of dorsal fin | 2.5 | 2.4 | 3.0 | |||
| Distance from anterior base of dorsal fin to “end of body” | 29.7 | 29.2 | [34.6] | |||
The measurements of these specimens of bairdii agree well together. The specimen of arnuxii appears to have had narrower flukes, shorter pectoral fin, and a rather higher dorsal fin, situated farther forward than in bairdii. Measurements of a larger number of specimens might show that some or all of these differences of proportion are elusive, but it will be observed that in the Wellington specimen of arnuxii, recorded by Doctor Haast, the breadth of the flukes is only 21 per cent of the total length. The pectoral fin is said to be only 19 inches long, or only 5.2 per cent of the total length, but the manner of taking the measurement is not mentioned.
As regards size, the largest specimen of B. arnuxii of which I find record is the type specimen. This was 32 feet long, and the skull 1,400 mm., or about 55 inches long. This appears to have been an adult male. The Centerville specimen of bairdii, which was an adult male, was about 41 feet long, and the skull 1,532 mm., or about 60 inches long, while the adult female from St. George Island was 40 feet 2 inches long and the skull 56 inches. Although the total length of the specimens of bairdii is so much greater, it will be observed that the length of the skull, while a little greater, absolutely fails to measure up to the proportions found in arnuxii. It might be suspected on this account that the external measurements of bairdii were exaggerated, but that such is not the case will appear from an examination of the measurements of vertebræ given on page 75. It is evident that the specimens of bairdii are far more massive in all parts of the skeleton than the specimen of arnuxii there cited. The same relations will be found upon comparing measurements of the specimen of arnuxii figured by Van Beneden and Gervais.[57] The truth appears to be that bairdii is a much larger species, but that the skull is considerably smaller relatively.
The skull of Berardius bairdii presents many characters by which it may be distinguished from that of arnuxii, whether adult or young. As compared with the latter, the rostrum is less massive at the base. The pterygoid has a rounded extension posteriorly and superiorly, so that the posterior portion of the upper border of the pterygoid sinus is convex, rather than nearly straight, as in arnuxii. The exoccipital is larger and broader distally below, and its external surface is plane or concave, rather than convex, as in arnuxii. The distal end of the zygomatic process is much more incurved. The nasal bones instead of presenting lateral extensions have nearly straight sides. The vomer is deeply emarginate at the base of the skull posteriorly where it rests against the presphenoid. The palatines extend scarcely or not at all in front of the pterygoids. The foregoing differences will readily be seen by comparing the figures on Pls. 26-29 with those of the type of B. arnuxii given in Van Beneden and Gervais’s Osteography, plate 23.
The following are dimensions of skulls of both species:
Dimensions of five skulls of Berardius bairdii (including the type) and of three skulls of B. arnuxii.
| A | B | C | D | E | F | G | H | |
|---|---|---|---|---|---|---|---|---|
| mm. | mm. | mm. | mm. | mm. | mm. | mm. | mm. | |
| Total length of skull | 1,372 | 1,392 | b1,174 | 1,524 | 1,423 | 1,378 | 1,062(?) | 1,474 |
| Height from vertex to inferior border of pterygoids | 533 | 494(?) | 493 | 563 | 544 | ... | ... | 575 |
| Breadth across middle of orbits | c625 | 684 | 577 | 766 | 682 | 662 | 530 | [716] |
| Breadth across postorbital processes | 686 | 748 | 606 | 808 | 722 | ... | 560 | [760] |
| Breadth across zygomatic processes | 671 | 748 | 584 | 750 | 675 | ... | 520 | [740] |
| Length of rostrum | 919 | 894 | 800 | 960 | 925 | 880 | 578+ | 1,025 |
| Breadth of rostrum at base | 399 | 414 | 378 | 475 | 420 | 428 | 310 | 429 |
| Breadth of rostrum at middled | 152 | 150 | 149 | 207 | 197 | 188 | ... | 223 |
| Length of premaxillæ | ... | .... | ... | ... | ... | ... | ... | ... |
| Breadth of premaxillæ at middled | 91 | 90 | 101 | 120 | 119 | 115 | ... | 125 |
| Greatest breadth of premaxillæ in front of nares | 208 | 210 | 189 | 235 | 217 | 238 | 187 | 239 |
| Greatest breadth of premaxillæ behind nares | ... | 246 | 193 | 215 | 195 | 181 | 165 | 197 |
| Distance from anterior end of premaxillæ posterior end of pterygoids (median) | 1,097 | 1,080 | 935 | 1,185 | 1,130 | ... | 720+ | 1,187 |
| Distance from anterior end of premaxillæ to anterior end of vomer | 345 | 264 | 252 | 276 | 270 | 260 | 115+ | 307 |
| Length of portion of vomer visible on palate | ... | 420 | 253± | 535 | 370 | 472 | 360± | 450 |
| Length of nasals (greatest, median, straight) | 132 | 162(?) | 134 | 135 | 118 | 135 | 98 | 142 |
| Breadth of nasals (greatest) | 102 | 180 | 125 | 119 | 97 | 105 | 90 | 105 |
| Breadth of anterior nares | 74 | 102 | 80 | 110 | 98 | 96 | 83 | 100 |
| Breadth of foramen magnum | 61 | ... | 72 | 85 | 82 | 84 | 83 | 71 |
| Breadth across occipital condyles | 191 | 213 | 186 | 261 | 228 | 240 | 195 | 235 |
| Breadth of each condyle | ... | ... | 75 | 123 | 104 | 108 | 83 | 98 |
| Height of each condyle | ... | ... | 135 | 193 | 171 | 168 | 142 | 178 |
| Length of mandible | e1,245 | 1,236 | ... | 1,334 | 1,289 | 1,282 | f883 | 1,360 |
| Length of symphysis | 310 | 294 | ... | 295 | 295 | 270 | f145 | 310 |
| Height at coronoid | 211 | 222 | ... | 271 | 230 | 223 | 175 | 245 |
| Distance from tip of jaw to center of first tooth | 34 | 45 | ... | 50 | 48 | 35 | f22 | 60 |
| Distance from tip of jaw to center of second tooth | 155 | 159 | ... | 200 | 182 | 165 | f87 | 195 |
The foregoing measurements indicate a considerable variation in proportions among the different individuals, but there appears to be nothing that can be fixed upon in this small series to distinguish the two species by dimensions alone.
The tympanic and periotic bones of B. bairdii (Pls. 34-37) present a number of characters by which they may be distinguished from those of B. arnuxii. While of about the same size in both species, the two bones when in the natural position, viewed from without, are nearly square rather than triangular in outline in B. bairdii, the superior border of the periotic being nearly parallel with the inferior border of the tympanic, and the anterior lobe of the periotic being turned down nearly at right angles with the rest of the bone. The periotic is shorter anteriorly than the tympanic in B. bairdii, while the reverse is true in B. arnuxii. In the former species the eustachian canal of the tympanic is wider, the distance between the outer and inner lips being greater. The involuted portion of the inner lip is shorter and differently shaped. The groove between the postero-inferior lobes is wider. The periotic beside having a much shorter anterior lobe than in B. arnuxii has also a smaller and smoother middle lobe, and the internal auditory meatus is smaller and more oblique. The dimensions of the bones in the Centerville beach skull, No. 49725, are as follows: Tympanic: greatest length, 62 mm.; greatest [70] breadth, 46; least breadth of eustachian canal, 17; height at sigmoid process, 47. Periotic: greatest length, 66; greatest breadth, 40; height at center of middle lobe, 35; length from tip of anterior lobe to anterior margin of internal meatus, 38.
Although all the specimens of Berardius bairdii are more or less incomplete, two or three of the mandibular teeth have been preserved in nearly every instance; namely, in the adult female from St. George Island, the left anterior and right and left posterior; in the immature male from the same island, both anterior teeth; in the Centerville beach specimen, the left anterior and right (?) posterior teeth; in the skull from Bering Island formerly regarded as the type, all four teeth; in the very young skull from Bering Island, the left anterior and posterior teeth.
Taken as a whole, these teeth are not larger than those found in the specimens of B. arnuxii thus far recorded, but in both species they vary so much on account of age, or for other reasons, that a comparison of dimensions is unsatisfactory. The dimensions are as follows:
Dimensions of teeth of Berardius arnuxii and B. bairdii.
| Species and locality. | Sex and age. | Length. | Large tooth. | Small tooth. | Remarks. | |||
| A | B | A | B | |||||
| B. arnuxii. | ft. | in. | mm. | mm. | mm. | mm. | ||
| Akaroa (Van Beneden. Type). | Male. | 32 | 0 | a90 | 90 | 66 | 40 | From figure.b |
| New Brighton (Haast and Flower). | Male(?). | 30 | 6 | a73 | 63 | 47 | 31 | From figure. |
| Port Nicholson (Knox and Hector). | (?) | 27 | 0 | a65 | 50 | (?) | (?) | From figure. |
| Locality unknown (Van Beneden and Gervais, pl. 21 bis). | (?) | (?) | 72 | 53 | 51 | 30 | From figure. | |
| B. bairdii. | ||||||||
| 49725—Centerville, California. | Male(?), adult. | 41 | ± | a83 | 65 | 53 | 28 | |
| 49726—St. George Island. | Female, adult. | 40 | 5 | c79 | 72 | 62 | 45 | |
| 49727—St. George Island. | Male, im. | 25 | 0 | a86 | 61 | ... | ... | |
| 142118—Bering Island | Young. | ... | ... | a50 | 37 | 31 | 31 | |
A description of the teeth of the different specimens of B. bairdii is subjoined.
No. 142118.—Bering Island; young (new born?). Anterior tooth conical, hollow, with thin walls. The lower half of the tooth is filled with a mass of bony pulp, which is separable. The tooth is widest at the base, and is without any constriction indicating the formation of a root. Outer and inner surfaces slightly convex, the latter with several distinct longitudinal furrows, which extend to the apex. The whole tooth has a thin coating of cement, except the tip, for a length of about 10 mm., which is more nearly white, and consists, presumably, of dentine. [71] The tooth is very symmetrical, but rather more convex externally. The apex is pointed, erect, and a little more convex externally than internally. (Pl. 39, figs. 1, 2.)
The posterior tooth is similar to the anterior one, but much shorter and more blunt, and the longitudinal furrows are about equally distinct externally and internally. The cement extends nearly to the apex, which latter is very short and is directed backward.
No. 49727.—St. George Island, Alaska; male, immature. Anterior teeth conical, acute, somewhat unsymmetrical, rather more convex externally than internally. The internal surface with a deep median longitudinal groove, and others less distinct on each side near the base. Apex slightly inclined forward and inward, convex externally, with a single longitudinal groove; nearly flat internally, with, or without, a groove. Base of tooth for about 17 mm. covered with longitudinal rugosities, indicating that the root was about to close. It is open, however, the walls of the tooth at the narrowest point being 8 mm. apart and the cavity filled with dense bony pulp. The anterior and posterior outlines of the teeth are irregular, being convex near the base, then slightly concave, and again convex near the apex. When in the natural position, these teeth protrude about 33 mm., or a little more than one-third their height, above the alveolus. (Pl. 39, figs. 3, 4.)
Posterior teeth lacking.
No. 49725.—Centerville beach, California; male (?), adult. Anterior tooth conical, with anterior and posterior margins as in the last. Apex considerably abraded and rounded off; not inclined inward or forward. Internal and external surfaces nearly equally convex, but the former with a broad median longitudinal groove. Root closed, the base of the tooth for a breadth of about 30 mm. covered with rounded rugosities. The inferior border slightly convex and the angles rounded off. When in the natural position, somewhat more than one-half of the tooth protrudes beyond the alveolus, and the tooth itself is inclined forward and outward. (Pl. 39, fig. 5.)
Posterior tooth quite irregular in form, but the portion above the rugose base or root conical. Inner surface flat and uneven. Outer surface convex and rather rugose. The cement covers the whole tooth thickly to within about 5 mm. of the apex, which latter is short, quite acute, and slightly directed inward. It is convex externally and nearly flat internally. The basal rugosity or root is conical, thicker than the rest of the tooth, and unsymmetrical, being somewhat directed backward. It shows no opening below. When in the natural position this tooth is strongly inclined forward and outward, and only the tip for a length of 22 mm. protrudes beyond the alveolus. (Pl. 39, fig. 6.)
No. 49726.—St. George Island, Alaska; female, adult. Anterior tooth conical, with the tip blunt, having been so much abraded that the dentine does not extend beyond the coating of cement. The tip measures 26 by 19 mm. The external and internal surfaces of the tooth are about equally convex and somewhat rugose without distinct furrows. The root is thicker than the remainder of the tooth and very rugose. It is entirely closed below, and the inferior outline is convex. Posterior tooth much compressed, conical above the root, nearly flat internally and slightly convex externally. Cement coating very thick and extending to within about 5 mm. of the dentine apex, which latter is acute and very slightly curved inward and [72] backward. The root is very unsymmetrical, the posterior portion being much longer than the anterior. The surface is very rugose, and there is no opening whatever below. The inferior border is convex, with an emargination near the center. (Pl. 39, figs. 7, 8.)
In the adult skull from Bering Island, which has been mounted and placed on exhibition, the teeth are fixed in the alveoli so that their entire length and the peculiarities of the basal portion can not be determined. In general form, however, they resemble those of the preceding specimen very closely. The anterior teeth are placed obliquely—that is, so that the anterior margins of the two teeth are nearer together than the posterior margins. The teeth are also somewhat inclined forward. The posterior teeth are strongly inclined forward and a little outward.
The anterior teeth are rather concave along the middle internally and convex externally. The portion above the alveoli is quite smooth.
The posterior teeth are moderately rugose above the alveoli. The whitish tips of denture are conical, compressed, and rather acute. They extend 6 mm. above the denture, and are 11 mm. long at their base, and 6 mm. thick.
The anterior teeth protrude about 45 mm. above the alveolus (internally); their base at the alveolus is from 73 to 76 mm. long, and from 33 to 35 mm. thick. The posterior teeth extend about 18 mm. above the alveoli (measured vertically from the alveolus), and the base of the visible portion (measured along the alveolus) is from 30 to 34 mm. long and from 18 to 20 mm. thick. These teeth have an antero-external angular enlargement of the cement, so that they are somewhat triangular in horizontal section. (Pl. 30, fig. 3; pl. 31, fig. 5.)
The data available are insufficient to enable one to determine satisfactorily whether the teeth differ materially in size in the two sexes, but it appears probable that they do not.
While the skeleton of Berardius bairdii (Pl. 42, fig. 4) resembles that of B. arnuxii very closely in most particulars, it presents differences which may properly be regarded as specific. The vertebral formula of B. arnuxii as given by Flower is as follows: C. 7, Th. 10, L. 12, Ca. 19 = 48.[58] The same formula is given for another specimen of B. arnuxii by Van Beneden and Gervais, except that the caudals are 17, two being apparently lacking.[59]
Doctor Hector, however, gives a different formula for a third specimen of this species, namely, C. 7, Th. 10, L. 13, Ca. 17 = 47. He remarks that “extreme care was taken to secure the whole of the small tail bones.”[60] The discrepancy here shown can not be accounted for at present, but, at all events, none of the formulas of B. arnuxii corresponds to that of B. bairdii, as derived from the three skeletons in the National Museum, namely, C. 7, Th. 11, L. 12, Ca. 16+ = 46+.
The number of thoracic vertebræ can be determined positively from the youngish male from St. George Island (Cat. No. 49727), in which ten pairs of ribs are present, together with one rib belonging to the eleventh pair. This last is much shorter [73] than the tenth pair, and there can be no doubt that it really belongs to a terminal pair. In this skeleton the transverse processes of the eleventh thoracic vertebra are thick at the free end like those of the tenth thoracic vertebra.
In the adult male from Centerville beach, California, only ten pairs of ribs are present, but as the tenth is quite as long as the ninth, there is little doubt that an eleventh pair was present originally. The eleventh thoracic vertebra, however, has transverse processes longer and more flattened at the free end than those of the tenth thoracic. It is possible, of course, that the real eleventh thoracic is lacking, and that this individual had thirteen lumbar vertebræ, but of this there is no positive evidence.
Only a few of the ribs accompany the skeleton of the adult female from St. George Island, Alaska (Cat. No. 49726), but there are eleven thoracic vertebræ, the transverse processes of the eleventh being short and thick, like those of the tenth, with a distinct facet for the rib at the free end. This facet, however, is directed obliquely backward and occupies only the posterior half of the free margin.
There is no doubt in my mind that the number of thoracic vertebræ in B. bairdii is normally 11 and in B. arnuxii, 10. This would ordinarily be of little importance, as in nearly all kinds of cetaceans a variation of one, or even two, in the number of thoracic and lumbar vertebræ in different individuals of the same species is commonly met with. In the present family, however, the number of thoracic vertebræ shows little variation, and as all known skeletons of B. bairdii have eleven thoracics and all known skeletons of arnuxii appear to have ten thoracics, it seems probable that this difference is specific. At all events, it is correlated with a difference in the form of the vertebræ themselves. As is well known, the transverse processes of the thoracics in this family undergo a sudden change of form and position near the end of the series, the elevated processes on the anterior thoracics being replaced on the posterior vertebræ by others at a lower level on the sides of the centra. This change takes place differently and on different vertebræ in the two species under consideration.
In B. arnuxii the eighth thoracic has no facet at the posterior end of the centrum for the articulation of the head of a ninth rib and no distinct transverse process, the tubercle of the rib articulating with a facet on the side of the metapophysis. In B. bairdii the eighth thoracic is similar, but there is a distinct facet at the posterior end of the centrum. (Pl. 32, fig. 1.)
In B. arnuxii the ninth thoracic has a very distinct transverse process on the side of the centrum, while in B. bairdii the ninth thoracic has a short, slender process attached to the side of the metapophysis and no facet at the posterior end of the centrum. (Pl. 32, fig. 1.)
In B. arnuxii the tenth thoracic is the second one having a distinct transverse process, and the latter is broad distally and has the articular facet on the posterior portion of the free margin. In B. bairdii the tenth thoracic is the first having a distinct transverse process on the side of the centrum. (Pl. 32, fig. 1.)
There are only ten thoracics in B. arnuxii, as already mentioned, but in B. bairdii there are eleven, and the eleventh is that which bears the second transverse process on the side of the centrum.
The foregoing differences amount to this: That in B. bairdii the commencement of the lower series of transverse processes is pushed back one vertebra, as compared with B. arnuxii, and that in the ninth thoracic of the former species, which corresponds to the eighth of the latter species, the metapophysis has a short process on the side for the articulation of the tubercle of the rib, instead of merely a sessile facet. Although in other genera of ziphioids these differences would perhaps be looked upon as individual, since they are constant here they may be considered specific, at least provisionally.
In B. bairdii the anterior border of the scapula is narrower than in B. arnuxii, the anterior ridge coming close to it and lying parallel with it. The acromion is directed more upward, so that the angle between it and the body of the scapula is more acute, and the process itself is rather more expanded distally. The coronoid is inclined a little more downward. The whole surface of the scapula is very uneven. (Pl. 33, fig. 2.)
The humerus is shorter than in B. arnuxii and broader distally, and much more recurved on the ulnar side. The ulna is much broader distally and its whole shape is different. (Pl. 33, figs. 3 and 4.)
As the skeleton of the typical form arnuxii has been described in considerable detail and accurately figured by Flower and by Van Beneden and Gervais, it is not considered necessary to give a complete description of that of bairdii in this place. The entire skeleton and many of the separate bones are figured in Pls. 42, 32, and 33. The phalanges are lacking altogether, or are incompletely represented, in the various skeletons of bairdii, and for that reason the phalangeal formula can not be given. The chevrons number ten in the skeleton from Centerville beach, California (Cat. No. 49725). Both Flower and Van Beneden and Gervais give nine chevrons as the number for the skeleton of arnuxii in the Hunterian Museum, London, but the latter authors have added a tenth in outline in the figure of the skeleton of that species which is in the Paris Museum. Ten are mentioned by Hector as the correct number for the skeleton of arnuxii from Wellington Harbor examined by him.[61]
The sternum of bairdii (Pl. 32, fig. 2) consists of five segments and does not offer characters by which to distinguish it from that of arnuxii. In the former species the first eight pairs of ribs possess distinct heads and tubercles; the tubercle is rudimentary in the ninth pair and absent in the tenth and eleventh.
The dimensions of the three skeletons of bairdii and of that of arnuxii described by Flower are as follows:
Dimensions of one skeleton of Berardius arnuxii and three skeletons of B. bairdii.
| B | C | D | E | |||||
|---|---|---|---|---|---|---|---|---|
| mm. | mm. | mm. | mm. | |||||
| Length of centra of seven cervicals (inferior) | 254 | 375 | 310 | 250 | ||||
| Atlas: | ||||||||
| Breadth | 292 | 362 | 341 | 280 | ||||
| Height | ... | 339 | 321 | 270 | ||||
| Fourth cervical: | ||||||||
| Greatest height | ... | a254 | a249 | a191 | ||||
| Greatest breadth | ... | b243 | b197 | b173 | ||||
| Length of centrum | 36 | 47 | 34 | 30 | ||||
| Seventh cervical: | ||||||||
| Greatest height | ... | 310 | 270 | 198 | ||||
| Greatest breadth | 241 | 257 | 235 | 177 | ||||
| Length of centrum | 46 | 58 | 49 | 42 | ||||
| First thoracic: | ||||||||
| Greatest height | ... | 391 | 390 | 255 | ||||
| Greatest breadth | ... | 310 | 290 | 240 | ||||
| Length of centrum | 58 | 84 | 68 | 51 | ||||
| Ninth thoracic: | ||||||||
| Greatest height | ... | 508 | 478 | 333 | ||||
| Greatest breadth | ... | 318 | c218 | 198 | ||||
| Length of centrum | 152 | 190 | 176 | 128 | ||||
| First lumbar: | ||||||||
| Greatest height | ... | 585 | 540 | 359 | ||||
| Greatest breadth | ... | 626 | 575 | 340 | ||||
| Length of centrum | 163 | 228 | 215 | 150 | ||||
| Sixth lumbar: | ||||||||
| Greatest height | ... | 713 | 642 | 427 | ||||
| Greatest breadth | ... | 590 | 572 | 362 | ||||
| Length of centrum | 206 | 273 | 243 | 172 | ||||
| First caudal: | ||||||||
| Greatest height | ... | 800 | d658 | 427 | ||||
| Greatest breadth | ... | 577 | d511 | 360 | ||||
| Length of centrum | 246 | 338 | d280 | 200 | ||||
| Ninth caudal: | ||||||||
| Greatest height | ... | 422 | 335 | 288 | ||||
| Greatest breadth | ... | 243 | 194 | 191 | ||||
| Length of centrum | 168 | 241 | 194 | 160 | ||||
| Eleventh caudal, length of centrum | 104 | 180 | 156 | 142 | ||||
| Length of scapula | 503 | 710 | 670 | 395 | ||||
| Height of scapula | 356 | 490 | 445 | 280 | ||||
| Length of humerus | 274 | ... | 340 | 248 | ||||
| Breadth of humerus at distal end | 109 | ... | 170 | 115 | ||||
| Length of radius | 295 | ... | a380 | 220 | ||||
| Breadth of radius at distal end | 84 | ... | 140 | 88 | ||||
| Length of ulna (incl. olecranon) | 323 | ... | ... | 241 | ||||
| Breadth of ulna at distal end | 79 | ... | ... | 71 | ||||
| Length of sternum | 1,143 | 1,455 | 1,530 | ... | ||||
| Breadth of first segment of sternum | 325 | 375 | 495 | ... | ||||
| Length of first rib (straight) | 457 | 543 | 505 | 323 | ||||
| Length of fifth rib (straight) | 991 | ... | ... | ... | ||||
| Length of tenth rib (straight) | 737 | ... | ... | ... | ||||
The National Museum has one skeleton of this well-known species, somewhat imperfect, which is labeled as having been obtained on the coast of Norway, and was received about the year 1875. Its catalogue number is 14499. This skeleton is about 19 feet long and has the following vertebral formula: C. 7; Th. 9; L. 9; Ca. 19 (+1?) = 44 (or 45). Eight chevrons are attached to the caudal vertebræ, and at least two more were present originally. The fifth thoracic vertebra has no facet on the centrum for the head of the sixth rib, but the latter articulates with a small facet on the side of the centrum of the sixth thoracic vertebra. The seventh thoracic has a well-developed transverse process on the side of the centrum. The ninth rib is shorter and more slender than the others. None of the transverse processes of the caudal vertebræ are perforated by foramina. These processes end on the eighth caudal, and the neural spines on the tenth caudal. The free ends of the neural spines of the thoracic and lumbar vertebræ are all more or less rounded. The pectoral limbs are incomplete.
So far as I am aware, only three examples of Hyperoödon have been taken on the coasts of the United States, as mentioned in the list on page 2. The skeleton of one of these (from North Dennis, Massachusetts) is in the Museum of Comparative Zoology, Cambridge, Massachusetts, and the skull of the second (from Newport, Rhode Island), which was a female, is in the Academy of Natural Sciences of Philadelphia.[62] This skull is represented in Pl. 32, fig. 3.
“Hyperodon semijunctus, sp. nov. The question whether a Hyperodon visits this side of the Atlantic, has at length been solved by the description which I have received through Dr. Alexander Wilcocks of this city, of a species taken in Charleston Harbor. This is well drawn up by Gabriel Manigault, who set up the specimen, which adorns the Charleston Museum. The points wherein it evidently differs from its congeners, the H. bidens and latifrons, are, first, the separation of the four posterior cervical vertebræ, the three anterior only being solidly anchylosed, instead of the seven, as in the known species, even in the young, according to Dr. J. E. Gray. Second, the possession of one or more pairs of ribs added to the flying series, and of two more vertebræ, including ten dorsal instead of nine. (Nine are given by Cuvier, Ossemens Fossiles, viii, 188; and Flower, Proc. Zool. Soc. Lond., 1864, 419, for the bidens.) Five ribs are connected with the sternum, of which the anterior articulates with the seventh cervical by its inferior head.
“I extract the following from Gabr. Manigault’s description:
“‘The superior maxillary bones are quite pointed in front and widen out toward the base of the snout. Their lateral edges become developed on each side into a prominent vertical ridge, which is slightly convex on the outer surface, and the reverse on the inner. These bones, after having widened out upon approaching the orbits, ascend vertically along with the occipital (the two together holding the frontal, which is quite perceptible, between them) and form at the back of the head a transverse ridge, which is quite high and very thick. From my not knowing by what name it was known, I did not satisfy myself concerning the presence of palatine tubercles. Another peculiarity of the head consists in the lower maxillary bones being provided each at its point with a single small and very sharp tooth. These were not noticed during the dissection, owing to their being too much imbedded in the integuments; they are now, however, quite visible. In the cavity of the skull is a septum of bone separating the cerebrum from the cerebellum (i. e., the tentorium). The first rib is very wide and short, and presents a marked contrast to the others. The sternum is quite flat and wide. The pectoral fins are small, and have been carefully preserved, with the various carpal and phalangeal bones kept together by their natural ligaments. As the skeleton stands, the fins consist only of the scapula, the humerus, the radius, and the ulna, with but few phalanges.
“‘The length of this specimen is between twelve and thirteen feet.’” (Proc. Acad. Nat. Sci. Phila., 1865, p. 15.)
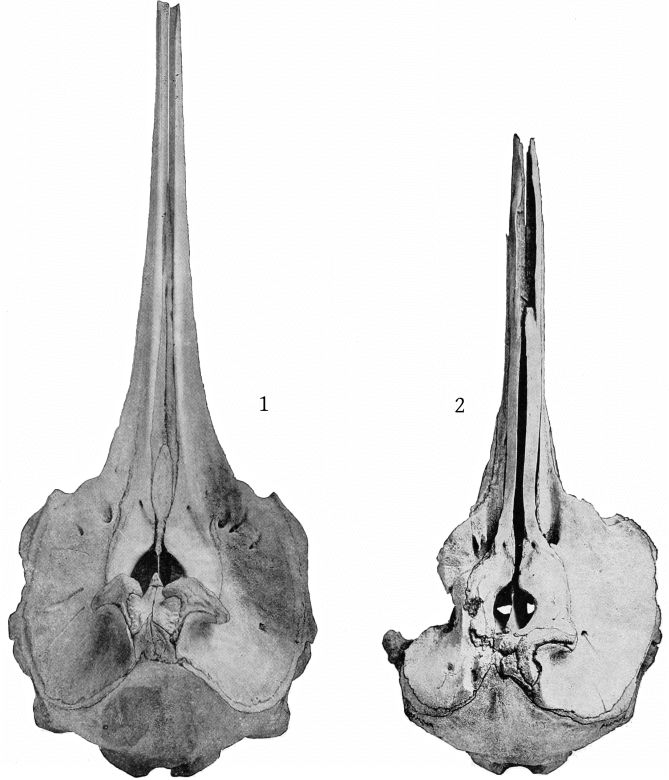
Fig. 1. Mesoplodon bidens. Skull. Nantucket, Mass. Mus. Comp. Zoölogy, No. 1727. Female, adult. Dorsal aspect. About ¼ nat. size.
Extremity of beak defective.
2. Mesoplodon densirostris? Skull. Annisquam, Mass. Female, young. Boston Society of Natural History. Dorsal aspect. ¼ nat. size.
Defective on the left side.
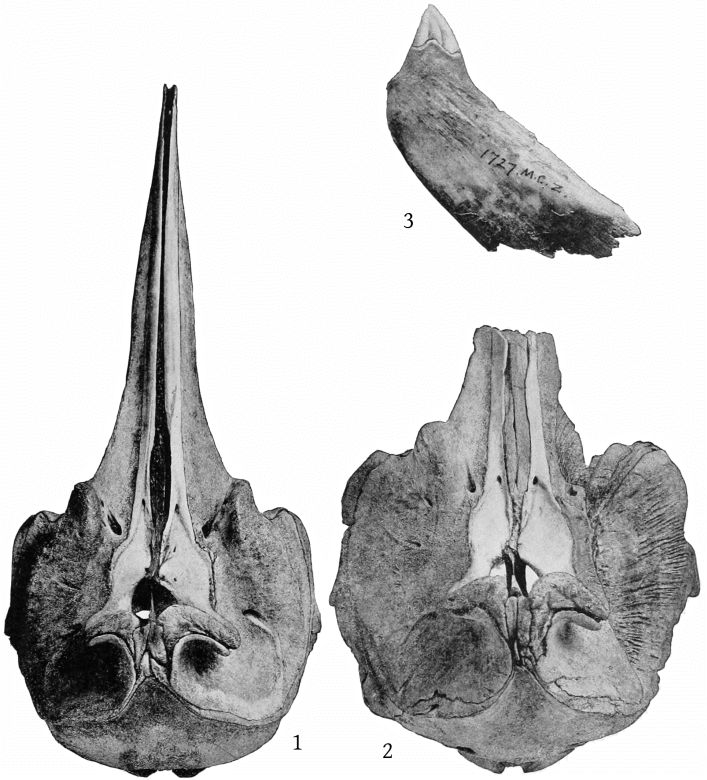
Fig. 1. Mesoplodon europæus. Skull. Atlantic City, New Jersey. Male, young. Cat. No. 23346, U.S.N.M. Dorsal aspect. ¼ nat. size.
2. Mesoplodon europæus. Skull. North Long Branch, New Jersey. Female, adult. Mus. Comp. Zoölogy. Dorsal aspect. ¼ nat. size.
Distal portion of beak lacking and right frontal region defective.
3. Mesoplodon bidens. Tooth. Nantucket, Mass. Mus. Comp. Zoöl., No. 1727. Nat. size.
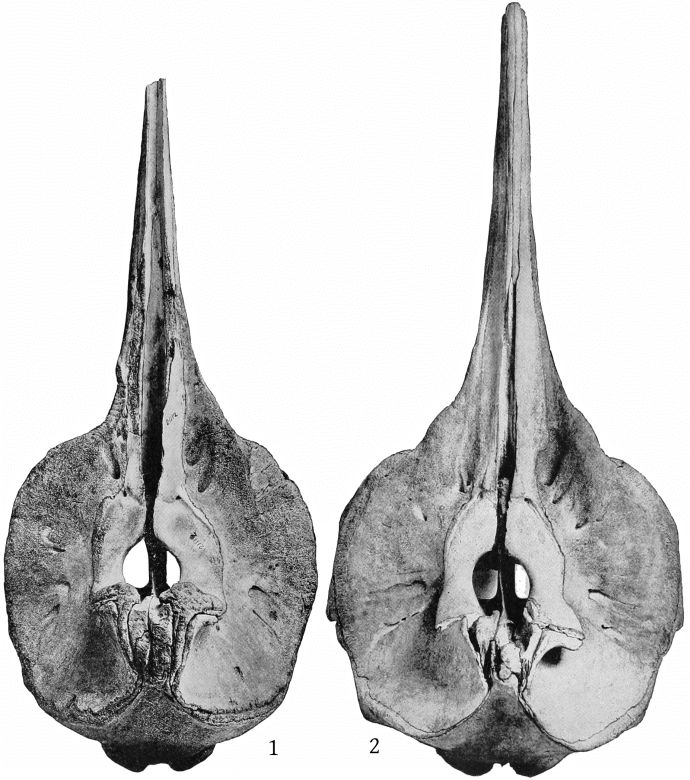
Fig. 1. Mesoplodon stejnegeri. Type-skull. Bering Island. Immature. Cat. No. 21112, U.S.N.M. Dorsal aspect. ¼ nat. size.
Edges abraded; distal end of beak defective.
2. Mesoplodon stejnegeri. Skull. Yaquina Bay, Oregon. Adult. Cat. No. 143132, U.S.N.M. Dorsal aspect. ¼ nat. size.
Proximal end of premaxillæ defective and right nasal lacking.
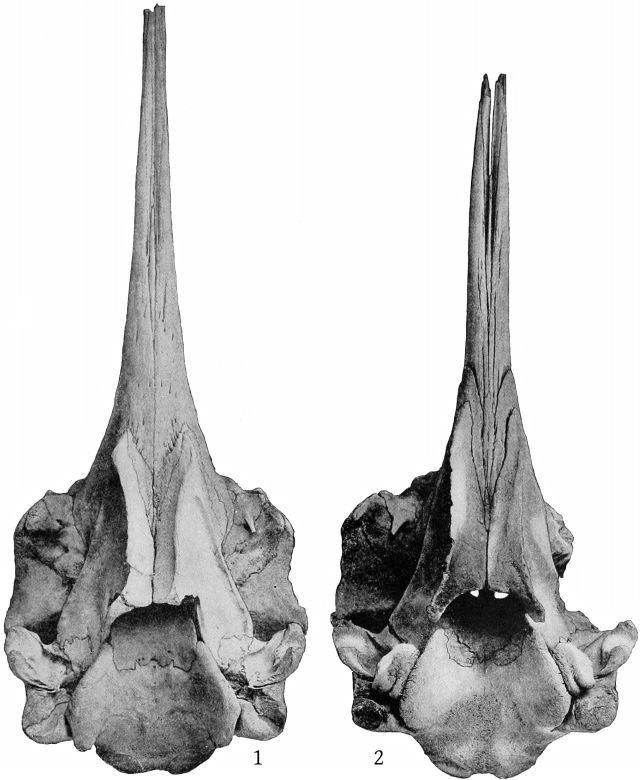
Fig. 1. Mesoplodon bidens. Skull. Nantucket, Mass. Female, adult. Mus. Comp. Zoöl. No. 1727. Ventral aspect. About ¼ nat. size. Tip of beak, left pterygoid, and malars defective.
2. Mesoplodon densirostris? Skull. Annisquam, Mass. Female, young. Boston Society of Natural History. Ventral aspect. ¼ nat. size.
Left frontal region defective.
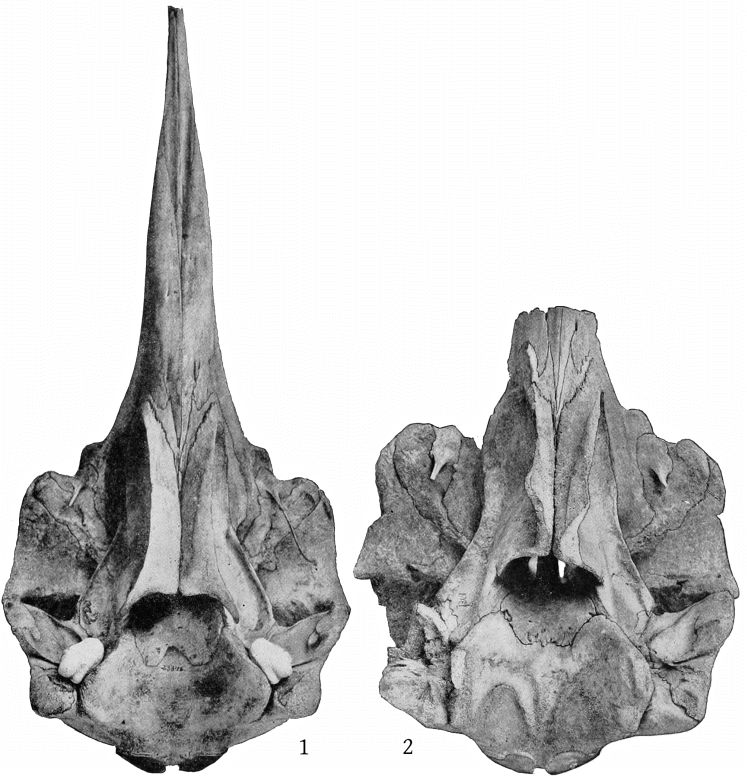
Fig. 1. Mesoplodon europæus. Skull. Atlantic City, New Jersey. Male, young. Cat. No. 23346, U.S.N.M. Ventral aspect. ¼ nat. size.
2. Mesoplodon europæus. Skull. North Long Branch, New Jersey. Female, adult. Mus. Comp. Zoölogy. Ventral aspect. ¼ nat. size.
Distal portion of beak lacking, pterygoids, malars, and left frontal and temporal regions defective.
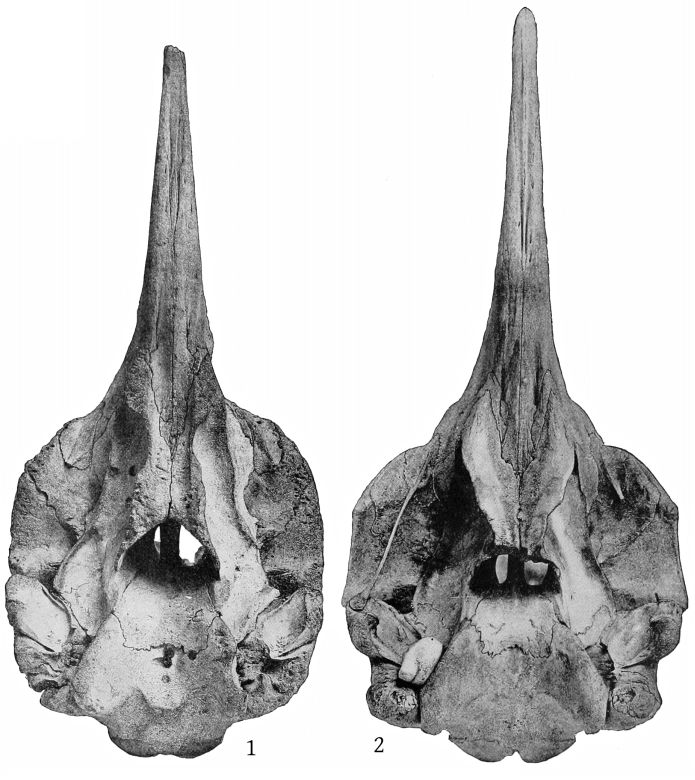
Fig. 1. Mesoplodon stejnegeri. Type-skull. Immature. Cat. No. 21112, U.S.N.M. Ventral aspect. ¼ nat. size.
Edges abraded; tip of beak, pterygoids, zygomatic processes, etc., defective.
2. Mesoplodon stejnegeri. Skull. Adult. Cat. No. 143132, U.S.N.M. Ventral aspect. About ¼ nat. size.
Pterygoids and left malar defective.
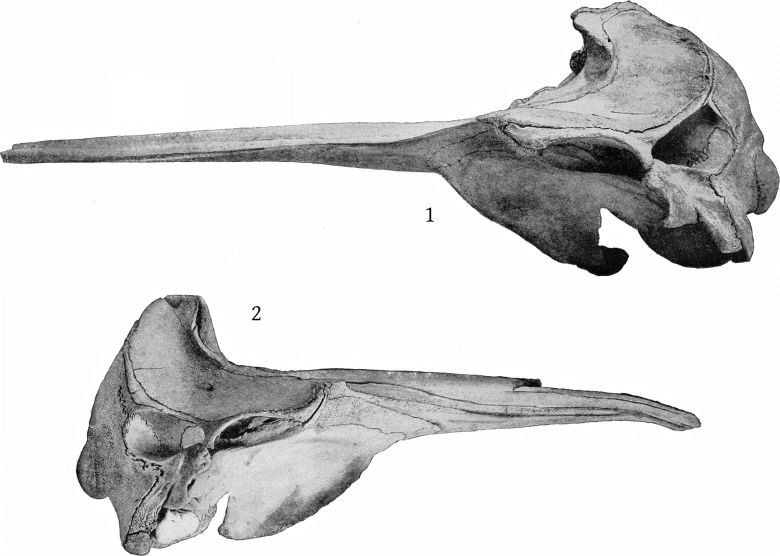
Fig. 1. Mesoplodon bidens. Skull. Nantucket, Massachusetts. Female, adult. Mus. Comp. Zoöl. No. 1727. Lateral aspect. ¼ nat. size.
Tip of beak, left pterygoid and malar defective.
2. Mesoplodon densirostris? Skull. Annisquam, Massachusetts. Female, young. Boston Soc. Nat. Hist. Lateral aspect. ¼ nat. size.
Distal portion of beak defective and warped.
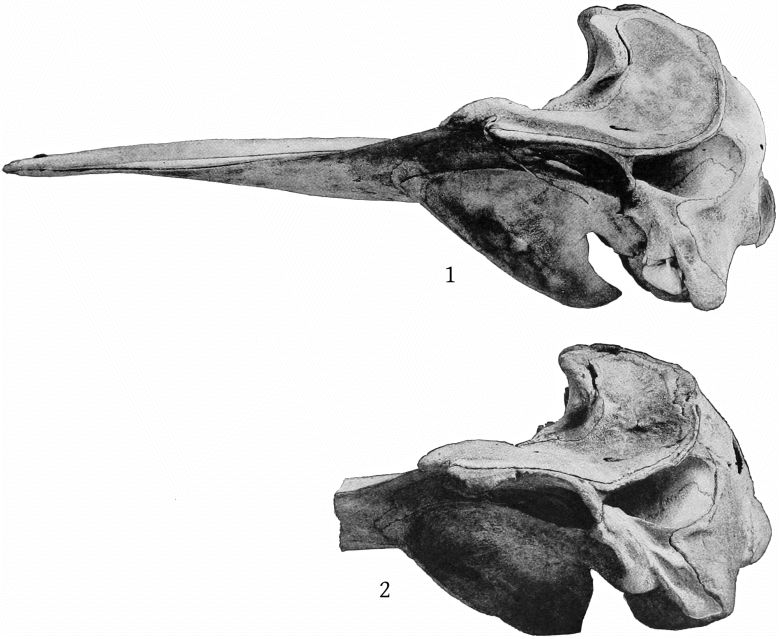
Fig. 1. Mesoplodon europæus. Skull. Atlantic City, New Jersey. Male, young. Cat. No. 23346, U.S.N.M. Lateral aspect. About ¼ nat. size.
2. Mesoplodon europæus. Skull. North Long Branch, New Jersey. Female, adult. Mus. Comp. Zoöl. Lateral aspect. About ¼ nat. size.
Distal portion of beak lacking.
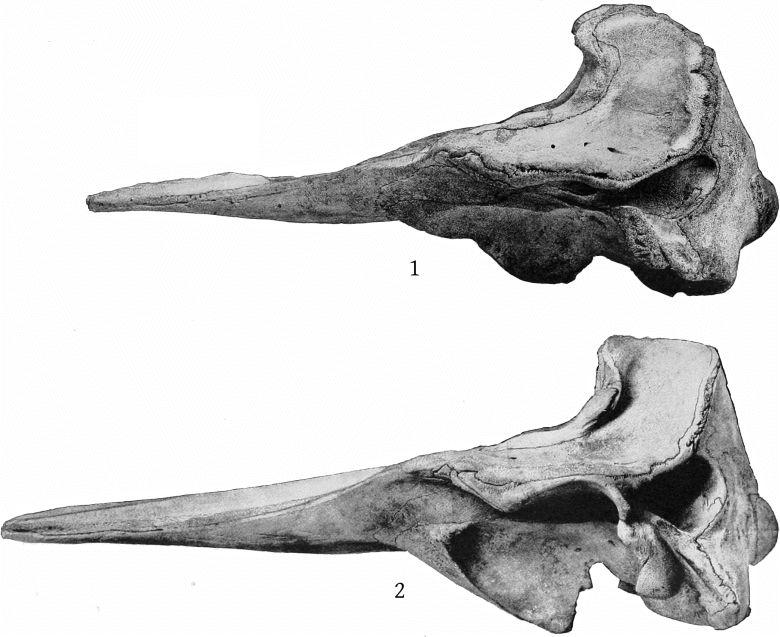
Fig. 1. Mesoplodon stejnegeri. Type-skull. Bering Island. Immature. Cat. No. 21112, U.S.N.M. Lateral aspect. About ¼ nat. size.
Premaxillæ, maxillæ, frontals, zygomatic process, etc., defective. On account of these defects and the immaturity of the individual the forward inclination of the supraoccipital is much greater than in the skull shown in fig. 2.
2. Mesoplodon stejnegeri. Skull. Yaquina Bay, Oregon. Adult. Cat. No. 143132, U.S.N.M. Lateral aspect. ¼ nat. size.
Proximal end of premaxillæ defective.
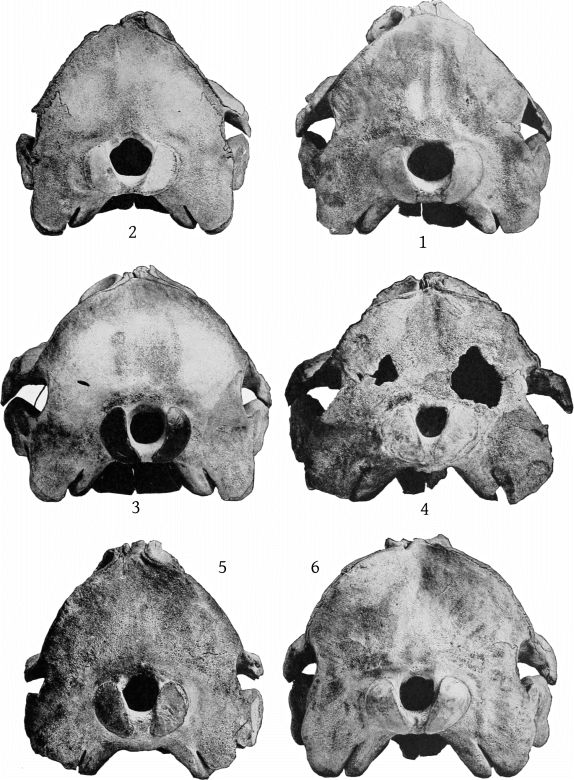
Skulls of Mesoplodon.
Fig. 1. Mesoplodon bidens. Nantucket, Massachusetts.
2. Mesoplodon densirostris? Annisquam, Massachusetts.
3. Mesoplodon europæus. Atlantic City, New Jersey.
4. Mesoplodon europæus. North Long Branch, New Jersey.
5. Mesoplodon stejnegeri. Type-skull. Bering Island.
6. Mesoplodon stejnegeri. Yaquina Bay, Oregon.
Posterior aspect. All figures ¼ nat. size.
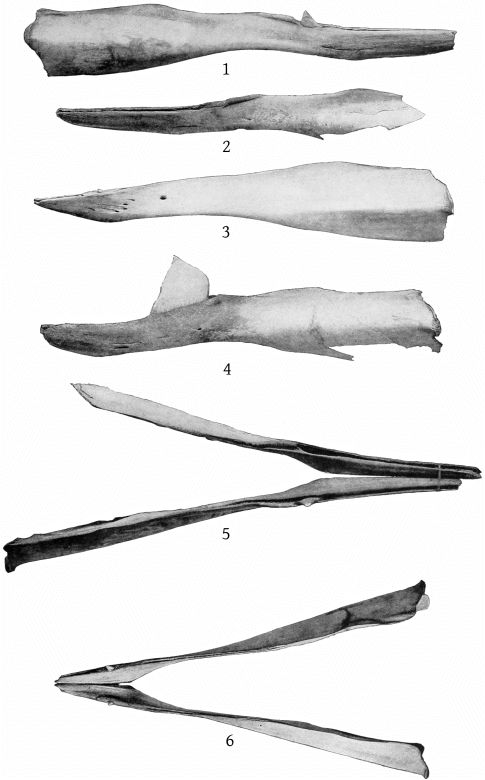
Mandibles of Mesoplodon.
Figs. 1, 2, and 5. Mesoplodon bidens. Nantucket, Massachusetts.
3 and 6. Mesoplodon europæus. Atlantic City, New Jersey.
4. Mesoplodon stejnegeri. Yaquina Bay, Oregon.
All figures ⅕ nat. size.
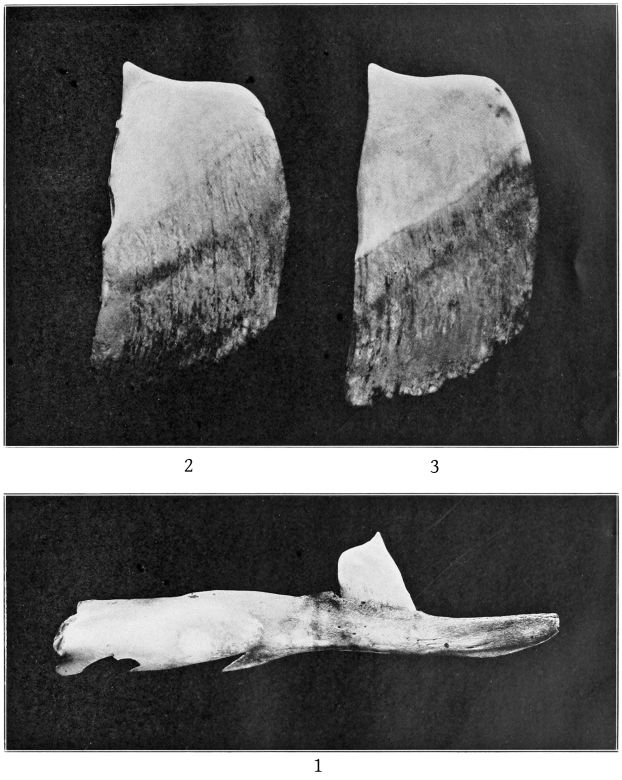
Fig. 1. Mesoplodon stejnegeri. Yaquina Bay, Oregon. Mandible and tooth. ¼ nat. size.
2. The same. Left mandibular tooth. Outer surface.
3. The same. Right mandibular tooth. Inner surface.
All figures a little more than ⅗ nat. size.
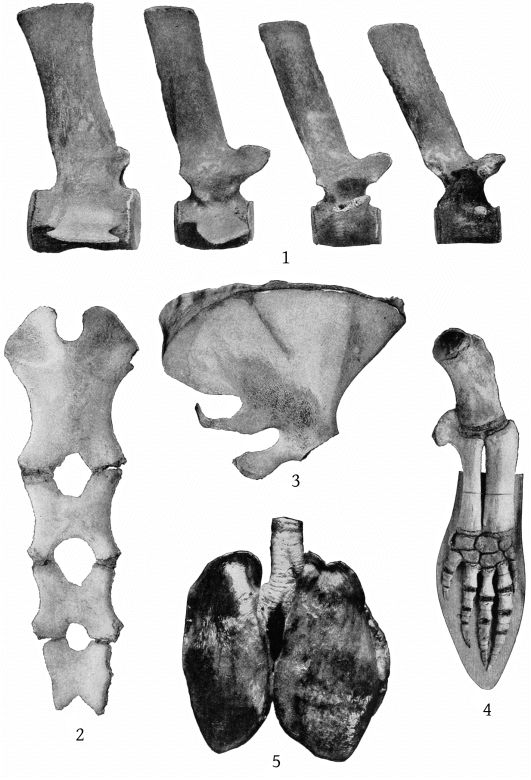
Mesoplodon europæus. Atlantic City, New Jersey. Cat. No. 23346, U.S.N.M.
Fig. 1. Vertebræ, from right to left as follows: 7th thoracic, 8th thoracic, 1st lumbar, 1st caudal. Scale, 1/3.7 nat. size.
2. Sternum. Anterior aspect.
3. Left scapula. External surface. Scale 1/3.6 nat. size.
4. Right pectoral limb. External surface. Scale 1/3.7 nat. size.
5. Lungs. Dorsal aspect. About 1/8 nat. size.

Ziphius cavirostris.
Fig. 1. Skull. (Type of Ziphius semijunctus (Cope).) Charleston, South Carolina. Female, young. Cat. No. 21975, U.S.N.M. Dorsal aspect. ⅙ nat. size.
Tip of beak slightly defective.
2. Skull. Barnegat City, New Jersey. Female, adult. Cat. No. 20971, U.S.N.M. Dorsal aspect. ⅙ nat. size.

Ziphius cavirostris.
Fig. 1. Skull. Bering Island. (Topotype of Ziphius grebnitzkii.) Female (?), adult, Cat. No. 22069, U.S.N.M. Dorsal aspect. ⅙ nat. size.
2. Skull. Bering Island. (Topotype of Ziphius grebnitzkii.) Dorsal aspect. Cat. No. 21246. ⅙ nat. size.
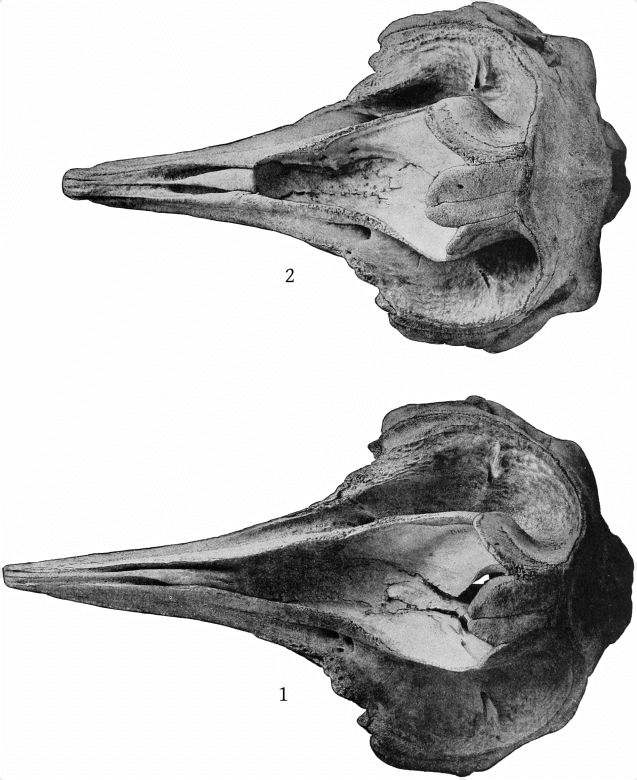
Ziphius cavirostris.
Fig. 1. Skull. (Type of Ziphius grebnitzkii Stejneger.) Bering Island. Male (?). Cat. No. 20993, U.S.N.M. Dorsal aspect. ⅙ nat. size.
2. Skull. (Topotype of Ziphius grebnitzkii.) Bering Island. Adult. Cat. No. 21245, U.S.N.M. Dorsal aspect. ⅙ nat. size.
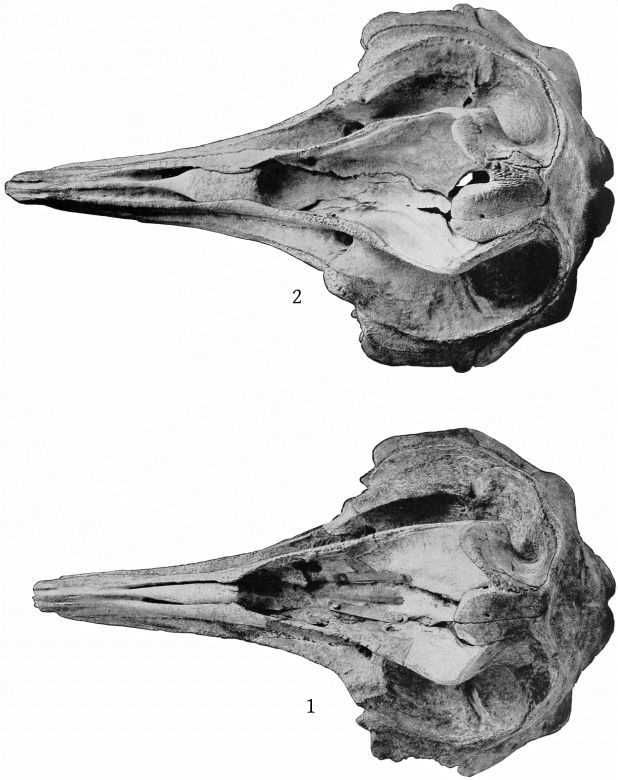
Ziphius cavirostris.
Fig. 1. Skull. (Topotype of Ziphius grebnitzkii.) Bering Island. Male (?), adult. Cat. No. 21248, U.S.N.M. Dorsal aspect. ⅙ nat. size.
2. Skull. Newport, Rhode Island. Male, adult. Cat. No. 49599, U.S.N.M. Dorsal aspect. ⅙ nat. size.

Ziphius cavirostris.
Fig. 1. Skull. (Type of Ziphius semijunctus (Cope).) Charleston, South Carolina. Ventral aspect. ⅙ nat. size.
2. Skull. Barnegat City, New Jersey. Ventral aspect. ⅙ nat. size.

Ziphius cavirostris.
Fig. 1. Skull. (Type of Ziphius grebnitzkii.) Bering Island. Cat. No. 20993, U.S.N.M. Ventral aspect. ⅙ nat. size.
2. Skull. Newport, Rhode Island. Ventral aspect. ⅙ nat. size.
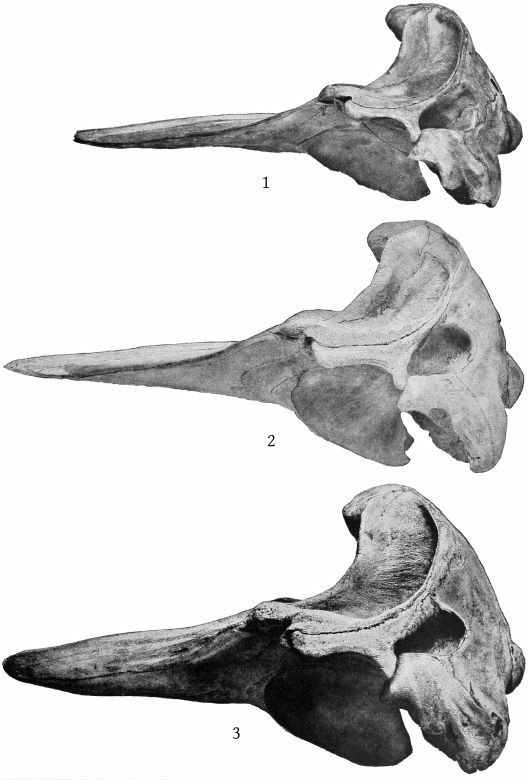
Ziphius cavirostris.
Fig. 1. Skull. (Type of Ziphius semijunctus (Cope).) Charleston, South Carolina. Cat. No. 21975, U.S.N.M. Lateral aspect. ⅙ nat. size.
2. Skull. Barnegat City, New Jersey. Lateral aspect. ⅙ nat. size.
3. Skull. (Type of Ziphius grebnitzkii Stejneger.) Bering Island. Cat. No. 20993, U.S.N.M. Lateral aspect. ⅙ nat. size.
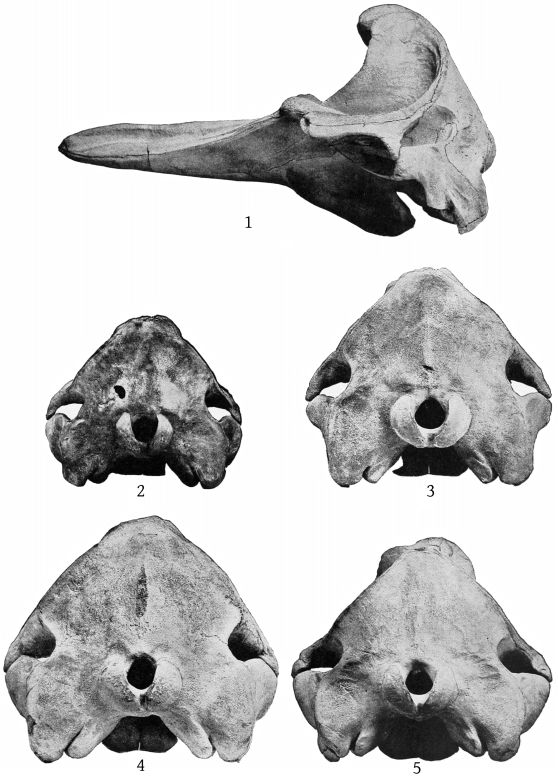
Ziphius cavirostris.
Fig. 1. Skull. Newport, Rhode Island. Lateral aspect. 1/7 nat. size.
2. Skull. (Type of Ziphius semijunctus (Cope).) Charleston, South Carolina. Posterior aspect. 1/7 nat. size.
3. Skull. Barnegat City, New Jersey. Posterior aspect. 1/7 nat. size.
4. Skull. (Topotype of Ziphius grebnitzkii Stejneger.) Posterior aspect. 1/7 nat. size.
5. Skull. Newport, Rhode Island. Posterior aspect. 1/7 nat. size.

Mandibles of Ziphius cavirostris.
Fig. 1. Charleston, South Carolina. (Type of Z. semijunctus (Cope).)
2. Newport, Rhode Island.
3. Bering Island. Cat. No. 22069, U.S.N.M.
4. Bering Island. Cat. No. 21248, U.S.N.M.
All figures about ⅕ nat. size.

Mandibles of Ziphius cavirostris.
Fig. 1. Bering Island. (Type of Ziphius grebnitzkii Stejneger.) Cat. No. 20993, U.S.N.M. About ⅕ nat. size.
2. Newport, Rhode Island. Symphysis. Dorsal aspect.
3. The same. Ventral aspect.
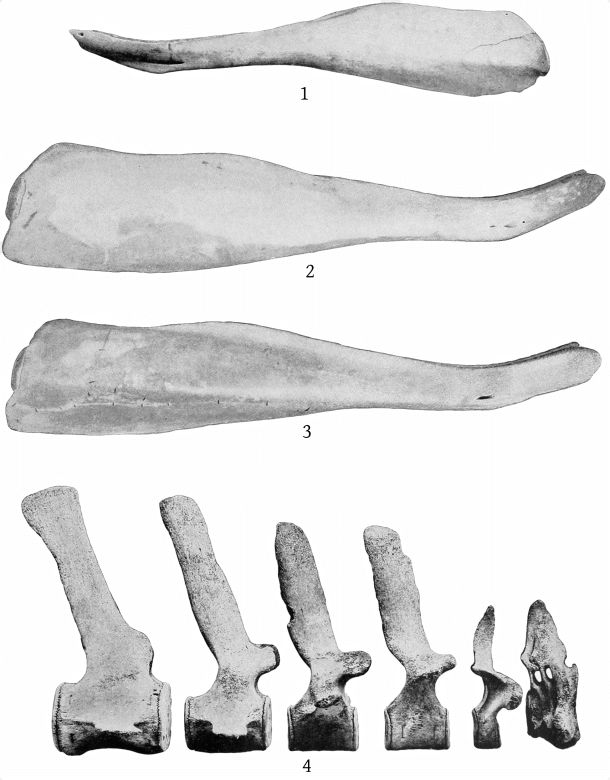
Mandibles and vertebræ of Ziphius cavirostris.
Fig. 1. (Type of Z. semijunctus (Cope).) Charleston, South Carolina. Cat. No. 21975, U.S.N.M. ⅕ nat. size.
2. (Type of Z. grebnitzkii Stejneger.) Bering Island. Cat. No. 20993, U.S.N.M. About ⅕ nat. size.
3. Barnegat, New Jersey. About ⅕ nat. size.
4. Vertebræ. (Type of Z. semijunctus (Cope).) From right to left, as follows: 1-3 cervicals, 1st thoracic, 7th thoracic, 8th thoracic, 1st lumbar, 1st caudal. About ¼ nat. size.
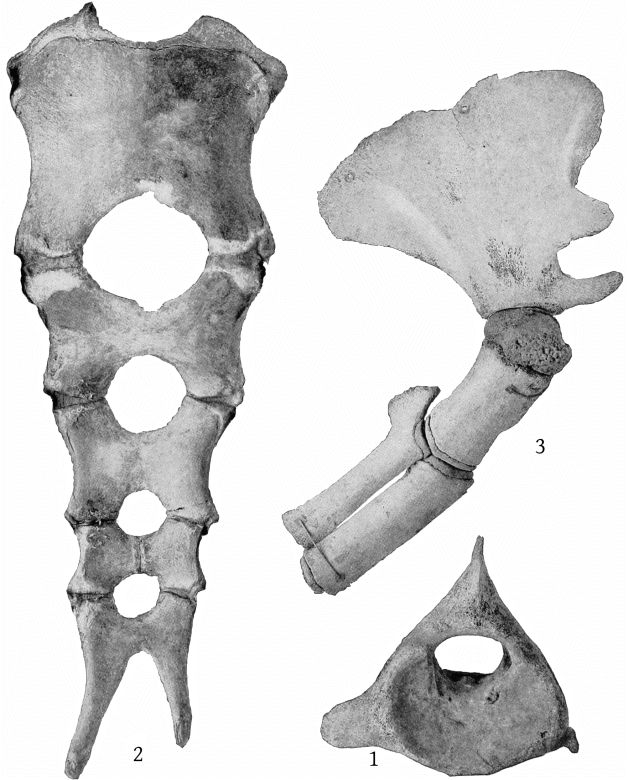
Ziphius cavirostris (Type of Z. semijunctus (Cope).)
Fig. 1. Atlas. Anterior surface. Defective on left side.
2. Sternum. Ventral aspect.
3. Right pectoral limb. Scapula somewhat defective.
About ࡩ nat. size.
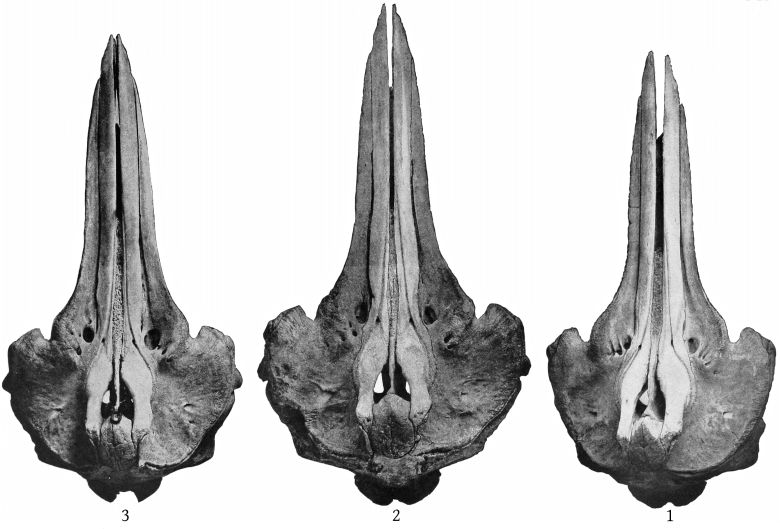
Berardius bairdii.
Fig. 1. Type-skull. Bering Island. Immature. Cat. No. 20992, U.S.N.M. Dorsal aspect. About 1/10 nat. size.
Frontals and zygomatic processes somewhat defective.
2. Skull. St. George Island, Pribilof Group, Alaska. Female, adult. Cat. No. 49726, U.S.N.M. Dorsal aspect. About 1/10 nat. size.
3. Skull. Centerville, California. Male (?), adult. Cat. No. 49725, U.S.N.M. Dorsal aspect.
All figs. about 1/10 nat. size.
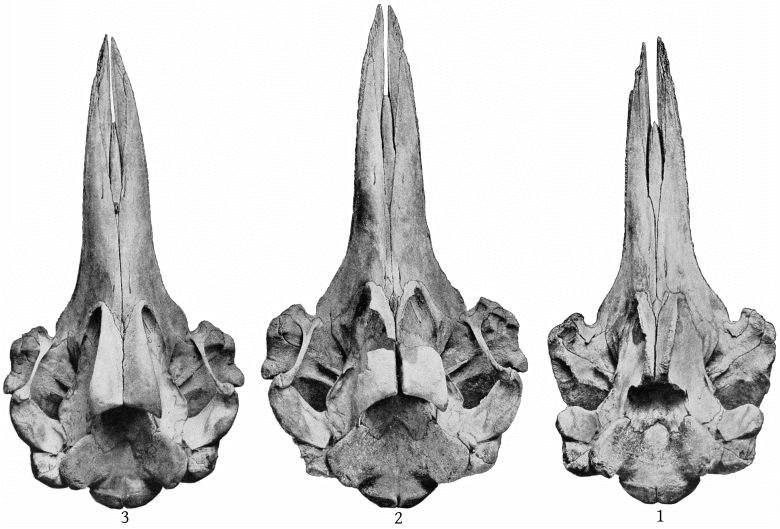
Berardius bairdii.
Fig. 1. Type-skull. Bering Island. Immature. Cat. No. 20992, U.S.N.M. Ventral aspect.
2. St. George Island, Alaska. Female, adult. Cat. No. 49726, U.S.N.M. Ventral aspect.
3. Centerville, California. Male (?), adult. Cat. No. 49725, U.S.N.M. Ventral aspect.
All figs. about 1/10 nat. size.

Berardius bairdii.
Fig. 1. Skull. St. George Island, Alaska. Female, adult. Cat. No. 49726, U.S.N.M. Lateral aspect.
2. Skull. Centerville, California. Male (?), adult. Cat. No. 49725, U.S.N.M. Lateral aspect.
3. The same skull. Posterior aspect.
4. Type-skull. Bering Island. Cat. No. 20992, U.S.N.M. Posterior aspect.
All figs. about 1/10 nat. size.
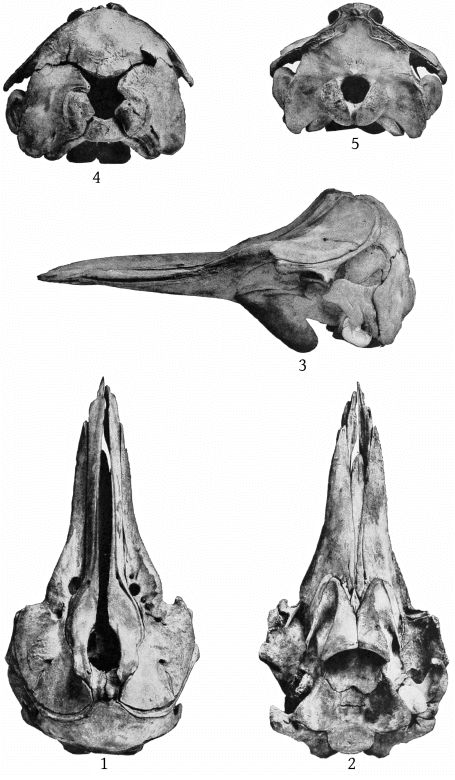
Berardius bairdii.
Figs. 1-4. Bering Island. Young. Cat. No. 142118, U.S.N.M.
5. Skull. St. George Island, Alaska. Male, immature. Cat. No. 29727, U.S.N.M. Posterior aspect. 1/10 nat. size.
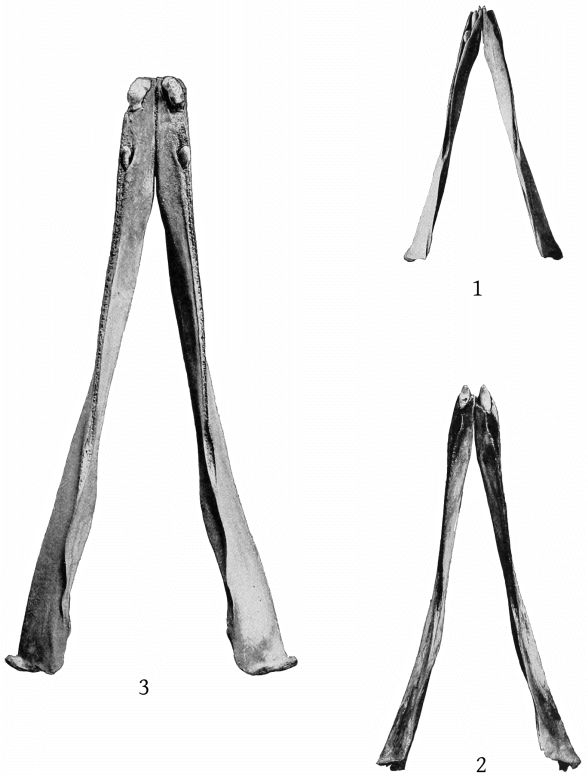
Mandibles of Berardius bairdii.
Fig. 1. Bering Island. Young. Cat. No. 142118, U.S.N.M.
2. St. George Island, Alaska. Male, immature. Cat. No. 49727, U.S.N.M.
3. Bering Island. Adult. (From mounted skull.)
Dorsal aspect, 1/10 nat. size.
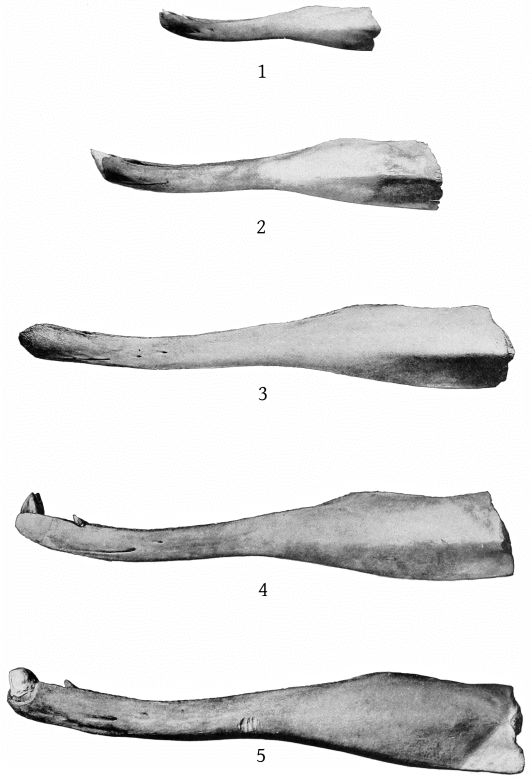
Mandibles of Berardius bairdii.
Fig. 1. Bering Island. Young. Cat. No. 142118, U.S.N.M.
2. St. George Island, Alaska. Male, immature. Cat. No. 49727, U.S.N.M.
3. Bering Island. (From type-skull.) Immature. Cat. No. 20992, U.S.N.M.
4. Centerville, California. Male (?), adult. Cat. No. 49725, U.S.N.M.
5. Bering Island. Adult. (From mounted skull.)
Lateral aspect. 1/10 nat. size.
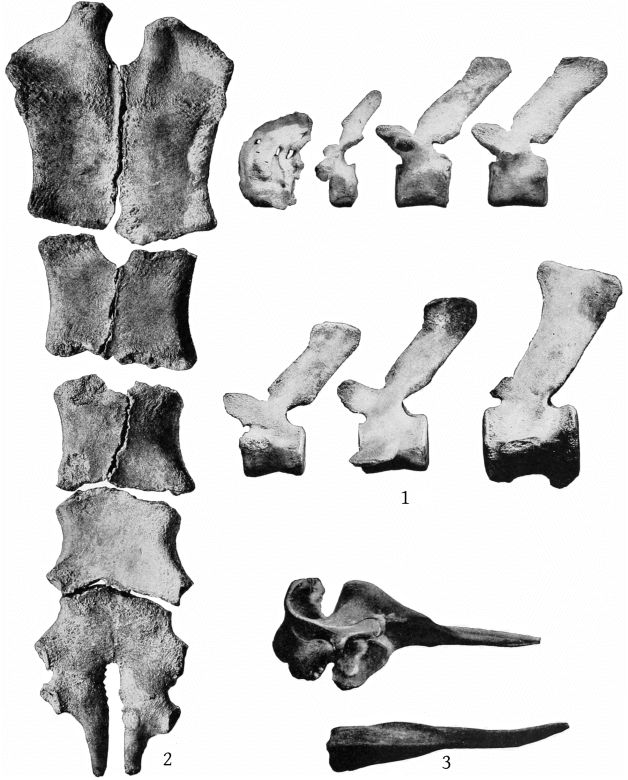
Berardius bairdii.
Fig. 1. Vertebræ. St. George Island, Alaska. Female, adult. Cat. No. 49726, U.S.N.M. The vertebræ from left to right are as follows: 1-3 cervicals, 1st thoracic, 8th thoracic, 9th thoracic, 10th thoracic, 1st lumbar, 1st caudal.
2. The same specimen. Sternum. Ventral aspect. About 1/7 nat. size.
3. Hyperoödon ampullatus. Newport, Rhode Island. Acad. Nat. Sci. Philadelphia.
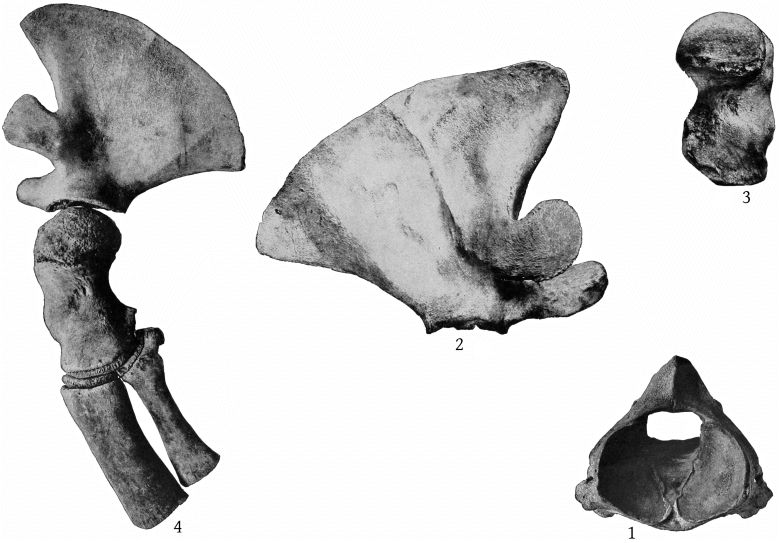
Fig. 1. Berardius bairdii. Atlas. St. George Island, Alaska. Female, adult. Cat. No. 49726, U.S.N.M. Anterior surface. 1/7 nat. size.
2. The same specimen. Right scapula. 1/7 nat. size.
3. The same specimen. Humerus. 1/7 nat. size.
4. Berardius bairdii. St. George Island, Alaska. Left pectoral limb. Cat. No. 49727. Male, immature. ⅕ nat. size.

Tympanic bones of Mesoplodon, Ziphius, and Berardius.
Fig. 1. Mesoplodon densirostris (?). Annisquam, Massachusetts.
2. Mesoplodon stejnegeri. Yaquina Bay, Oregon.
3. Ziphius cavirostris. (Type of Z. semijunctus (Cope).) Charleston, South Carolina.
4. Ziphius cavirostris. (Type of Z. grebnitzkii Stejneger.) Bering Island.
5. Ziphius cavirostris. Barnegat City, New Jersey.
6. Ziphius cavirostris. Newport, Rhode Island.
7. Berardius bairdii. Centerville, California.
Ventral aspect. Nat. size.
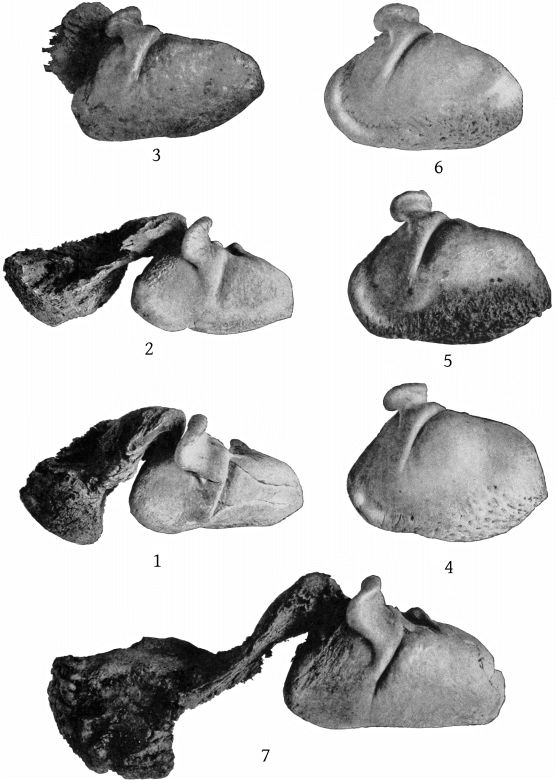
Tympanic bones of Mesoplodon, Ziphius, and Berardius.
Fig. 1. Mesoplodon densirostris (?). Annisquam, Massachusetts.
2. Mesoplodon stejnegeri. Yaquina Bay, Oregon.
3. Ziphius cavirostris. (Type of Z. semijunctus (Cope).) Charleston, South Carolina.
4. Ziphius cavirostris. (Type of Z. grebnitzkii Stejneger). Bering Island.
5. Ziphius cavirostris. Barnegat City, New Jersey.
6. Ziphius cavirostris. Newport, Rhode Island.
7. Berardius bairdii. Centerville, California.
External surface. Nat. size.
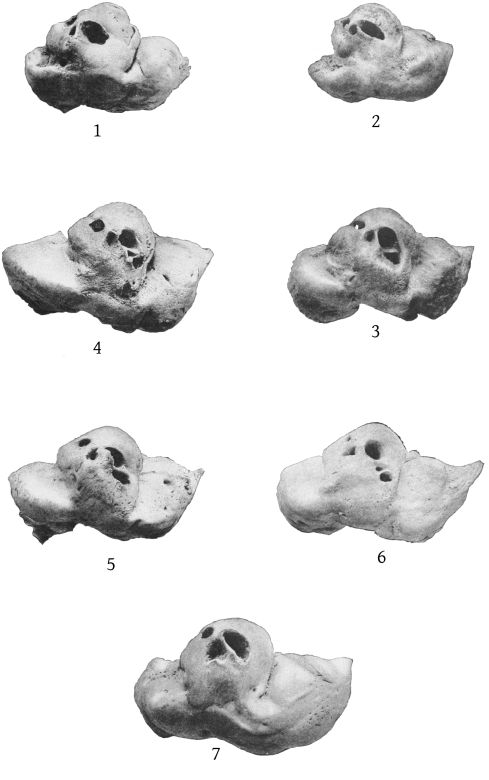
Right periotic bones of Mesoplodon, Ziphius, and Berardius.
Fig. 1. Mesoplodon densirostris. (?) Annisquam, Massachusetts.
2. Mesoplodon stejnegeri. Yaquina Bay, Oregon.
3. Ziphius cavirostris. (Type of Z. semijunctus (Cope).) Charleston, South Carolina.
4. Ziphius cavirostris. (Type of Z. grebnitzkii Stejneger.) Bering Island.
5. Ziphius cavirostris. Barnegat City, New Jersey.
6. Ziphius cavirostris. Newport, Rhode Island.
7. Berardius bairdii. Centerville, California.
Inner aspect. Nat. size.
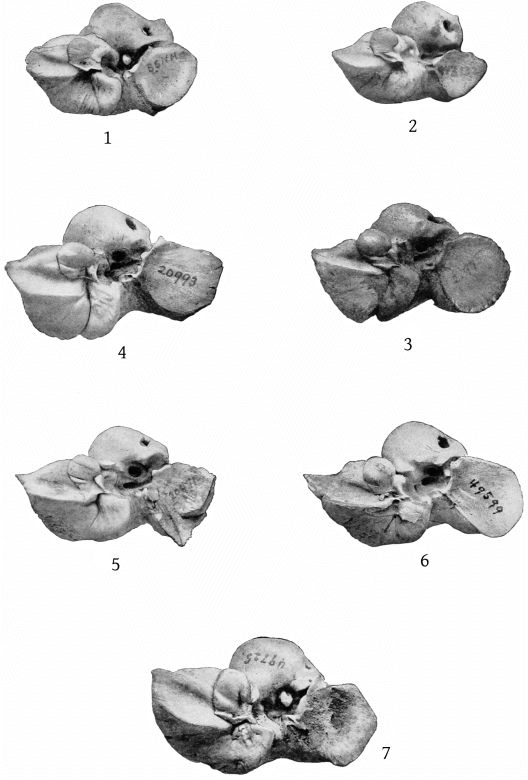
Right periotic bones of Mesoplodon, Ziphius, and Berardius.
Fig. 1. Mesoplodon densirostris. (?) Annisquam, Massachusetts.
2. Mesoplodon stejnegeri. Yaquina Bay, Oregon.
3. Ziphius cavirostris. (Type of Z. semijunctus (Cope).) Charleston, South Carolina.
4. Ziphius cavirostris. (Type of Z. grebnitzkii Stejneger). Bering Island.
5. Ziphius cavirostris. Barnegat City, New Jersey.
6. Ziphius cavirostris. Newport, Rhode Island.
7. Berardius bairdii. Centerville, California.
Outer aspect. Nat. size.
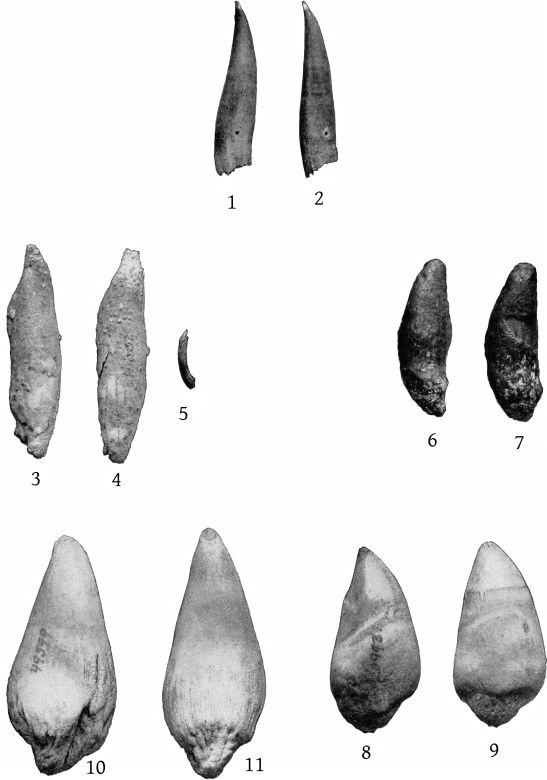
Teeth of Ziphius cavirostris.
Fig. 1. Type of Z. semijunctus (Cope). Charleston, South Carolina. Cat. No. 21112, U.S.N.M. Left tooth. Inner surface.
2. The same. Right tooth. Outer surface.
3-4. Barnegat City, New Jersey. The two large teeth.
5. The same. One of the rudimentary teeth.
6. Topotype of Z. grebnitzkii. Cat. No. 22069, U.S.N.M. Bering Island. Left tooth. Outer surface.
7. The same. Right tooth. Inner surface.
8. Type of Z. grebnitzkii Stejneger. Cat. No. 20993, U.S.N.M. Bering Island. Left tooth. Inner surface.
9. The same. Right tooth. Outer surface.
10. Newport, Rhode Island. Cat. No. 49599, U.S.N.M. Left tooth. Inner surface.
11. The same. Right tooth. Outer surface.

Teeth of Berardius bairdii.
Fig. 1. Bering Island. Young. Cat. No. 142118, U.S.N.M. Left anterior tooth. Inner surface.
2. The same. Left posterior tooth. Inner surface.
3. St. George Island, Alaska. Male, immature. Cat. No. 49727, U.S.N.M. Right anterior tooth. Inner surface.
4. The same. Left anterior tooth. Outer surface.
5. Centerville, California. Male, adult. Cat. No. 49725, U.S.N.M. Left anterior tooth. Inner surface.
6. The same. Right posterior tooth. Outer surface.
7. St. George Island, Alaska. Female, adult. Cat. No. 49726. Left anterior tooth. Inner surface.
8. The same. Right posterior tooth. Outer surface.
9. The same. Left posterior tooth. Inner surface.
All figures natural size.
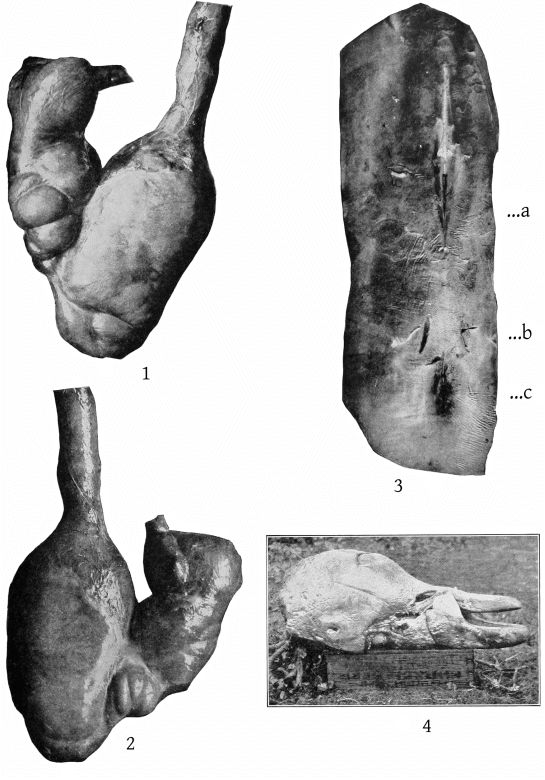
Fig. 1. Stomach of Mesoplodon europæus. Atlantic City, New Jersey. Cat. No. 23346, U.S.N.M. Ventral aspect. About ¼ nat. size.
2. The same. Dorsal aspect. About ¼ nat. size.
3. The same. Perineum. a, penis. b, rudimentary mammary slits. c, anus. About ¼ nat. size.
4. Mesoplodon stejnegeri. Yaquina Bay, Oregon. Cat. No. 143132, U.S.N.M. Head, showing teeth in natural position.

Fig. 1. Mesoplodon europæus. Atlantic City, New Jersey. Male, young. Cat. No. 23346, U.S.N.M. Length 12½ feet.
2. The same. Dorsal aspect.
3. Ziphius cavirostris (?). Kiska Harbor, Alaska, 1904.
4. Ziphius cavirostris. Newport, Rhode Island. Male, adult. Length 20 feet 1 inch. Cat. No. 49599, U.S.N.M.

Fig. 1. Berardius bairdii. St. George Island, Alaska. Female, adult. Length 40 feet 2 inches. Cat. No. 49726, U.S.N.M.
Ventral aspect.
2, 3. Berardius bairdii. Centerville, California. Male (?), adult. Cat. No. 49725, U.S.N.M. Length about 41 feet. Head from in front and from below.
4. The same. Skeleton. About ⅙ nat. size.
The pectoral fin is modeled from another specimen. It is on the wrong side in this figure.