
Title: Fossils: A Story of the Rocks and Their Record of Prehistoric Life
Author: Harvey C. Markman
Illustrator: Mary Chilton Gray
Release date: July 22, 2017 [eBook #55168]
Most recently updated: October 23, 2024
Language: English
Credits: Produced by Stephen Hutcheson and the Online Distributed
Proofreading Team at http://www.pgdp.net
By Harvey C. Markman
Curator of Geology and Paleontology
Cover Design and Murals by
Mary Chilton Gray
DENVER MUSEUM OF
NATURAL HISTORY
Popular Series No. 3
Alfred M. Bailey, Editor
Third Edition, Reprinted
October 1, 1954

At Work on a Dinosaur Skeleton
In the recent growth of knowledge there has been rapid progress in two directions. The commercial exploitation of natural resources, being fundamental to modern civilization, attracts a liberal share of the talents and energies of workers trained for the industrial professions. A second trend has specialized in the further development of the sciences which are characteristic of our time. Such activities, in the natural history field, deal largely with the refinements of exact definition, nomenclature and classification, all of which means little or nothing to the layman who is otherwise engaged.
For the latter, however, there is a quality of interest which may be described as a wholesome curiosity about what has happened, how it happened, how we know it happened, and what it may signify to one who is neither industrialist nor scientist. This booklet is intended for the many who feel that there is more to be obtained from a natural history museum than an occasional glimpse of a bewildering “marvel.” In addition to being a guide to fossil exhibits it supplies parts of a great story which specimens alone can not relate.
All that is known of the extinct plants and animals which inhabited the earth before man began the practice of recording his observations has been obtained from a study of the rocks. The few possible exceptions to this rule, in which animal and plant remains have been preserved by freezing or drying, are so unusual as to be hardly worth mentioning.
Explanation of this is that plant and animal tissues quickly decay under ordinary conditions when life ceases. Unless protected from destructive agencies which are especially active at the surface of the ground, even the heavier bones of animals and the large trunks of fallen trees will soon crumble into shapeless masses. The usual method employed by nature to prepare a fossil specimen is so closely related to the natural process of rock making that a little knowledge of that subject will be necessary in order to know what fossils are and how they are preserved for so long a time.
It should be understood first that a fossil is some record, commonly preserved in rock, of a kind of plant or animal which no longer exists 6 as a living type. This, at least, is the ordinary sense of the word and more elaborate definitions are of small service to anyone. It may be necessary to add, however, that all things which have lived at any time are regarded as either plants or animals.
Nature’s way of producing rocks and fossils remains a mystery to many of us because we are so wrapped up with the importance of finding names for things and materials that we frequently neglect the consideration of sources and histories. Everyone knows a rock when he sees it in a large mass, but when he looks at sand, mud, dust, or soil, he seldom thinks of it as related in any way to rocks. Although the difference is almost entirely a matter of size, our use of words makes it seem unreasonable to speak of the finer particles as rock.
There can be no reality or meaning in the natural record for an individual who has failed to observe a few simple facts which involve changes going on in all parts of the world at the present time. With regard to rocks, it is supposed that what happens in our day also occurred under like circumstances ages ago. Anyone wishing to do so may see for himself that rock masses break down wherever they are exposed to the elements, that the larger pieces are reduced to smaller fragments, and that the final product is sand or dust.
He may also note that this finely ground material is being moved and sorted, by rain, wind, and streams, transported to lower levels and accumulated in great quantities wherever it finds a resting place. Along with it go sticks and leaves, bugs, shells, bones and carcasses of animals, some of which in time may become fossils. In large lakes and seas there is a steady distribution of such materials over broad areas, yesterday’s accumulation of sediments being buried by the contributions of today, the most recent of the settlings always resting upon older ones until something happens to disturb that arrangement.
Not so readily observed are other parts of the process, such as the consolidation of sands and muds into the firm sandstones and shales which we again recognize as rocks. Much of this requires more than the few score years of a human lifetime for its accomplishment, but many of us have seen muds become so solidified, by merely drying, that they could hardly be distinguished from prehistoric shales. It is to be noted also that some ancient fossils come from sandstones which are scarcely more rock-like than the loose sands of an ocean beach. Thus we learn that firm consolidation of rock-making materials is not always a sign of great antiquity, and that hardness of rock is not always essential to the preservation of imbedded plants and animals.

Rhinocerous Bones as Found in the Rock
This exhibit was taken from the famous fossil quarry at Agate, Nebraska. The fossilized remains are still partially imbedded in the sandstone which preserved them for millions of years.
The rocks themselves must explain the many things which have happened during the course of millions of years, and this they do remarkably well when carefully studied, for many of the factors involved in their histories leave characteristic marks. Changing climates, the draining of seas, the uplifting of mountain ranges, all have ways of registering their occurrence which are as convincing and reliable as anything ever written by man. Piece by piece the story has been patched together through the efforts of thousands of investigators. Parts of the narrative remain buried at inaccessible depths, and whole chapters, no doubt, have been destroyed by the same forces that composed this tremendous record of prehistoric times.
It would be a serious mistake to regard nature as divided into a number of distinct and independent schools of fossil making, each refusing to use the methods and devices of another. There are, however, certain features which stand out so prominently that a little classification becomes helpful. While this procedure brings out differences it should be understood that processes actually work together, several of them usually being involved in the production of any individual specimen.
(1) Impressions of animals and plants, or parts of these, are frequently left in soft sand or mud which later becomes converted into more durable rock. This type of fossils is represented by animal foot-prints and the imprints of leaves, flowers, insects, and like objects which may be mingled with the finely ground materials of the common sedimentary rocks.
(2) Parts of plants and animals may be gradually replaced by mineral matter with little or no change from original form and texture. Fossils of this class are said to be petrified or turned to stone. They are also known as replacements. The fleshy parts of animals do not petrify.
(3) Many animals among the invertebrates use mineral substances for protective or supporting structures. Small plants of various kinds follow a similar practice. These structures, being produced in stony materials, are readily converted into fossils. The shells of mollusks are the best known illustrations in this field, and all that is required for a shell to become a fossil is the extinction of the species of animal that produced it. Fossils of this type are extremely abundant.
(4) Preservative substances other than those which produce common rocks may be mentioned among fossil-making possibilities. Bones are known to have been preserved in asphalt, and insects in resins, but such cases are few in comparison with the products of other methods.
(5) In rare instances there has been preservation of extinct creatures by the process of drying or by refrigeration. Occasional mummies 9 are found with shriveled flesh and skin still in place, but better preservation of all tissues occurs when the temperature is quickly reduced below freezing point and held there without interruption. This can happen only in the colder parts of the earth and is always subject to climatic change. The effect of drying also may be undone at any time by a slight increase in the amount of moisture.
(6) Coal beds often produce fossils of an unusual sort. In the formation of coal, plant material gradually loses some of its more perishable substances but retains carbon which has better lasting qualities and slowly accumulates to produce the seams and beds that are mined. In the early stages of the process the original vegetation undergoes little change in appearance but eventually practically all of its character is lost. Many fossil leaves are found as thin layers of carbon, bedded in the clays which are commonly associated with coal deposits.
(7) Concretions, which are hardened lumps of mineral substances occurring commonly in sandstones and shales, are often mistaken for fossils because of their peculiar shapes. However, there are localities in which the mineral solutions have been concentrated and deposited around shells, leaves, seeds, or similar objects, thus producing an abundance of fossils which may be obtained by opening the concretions. Fossils of this type are well known from Mazon Creek and other districts in Illinois, Kansas, Colorado, and elsewhere.
Footprints need little explanation other than a consideration of the factors which make it possible for them to be preserved. The sand or mud must be neither too soft nor too hard to take the form of the foot and retain its shape when the foot is withdrawn. Then in some manner the impression must be protected while the rock-making process goes on. When such protection is obtained it is usually in the form of more mud and sand, deposited over the surface which received the impression. At a later time the covering may be separated from the lower part of the deposit, which serves as a mold, and if the separation be accomplished successfully a natural cast of the foot will be obtained as well as the mold in which it was produced. Since conditions for perfect work are not always present in a laboratory of this kind, it is not surprising that fossil footprints are very rare considering the number and variety of tracks left by wandering animals.
Impressions of leaves are explained in much the same way except that the leaf remains under its protective covering until it decays. Similar impressions may be obtained from the bodies of delicate invertebrate animals 10 but they are seldom preserved because of the softness of the tissues. The smaller fishes provide much better material for the production of fossils according to this method. While the fish is being flattened by the weight of surrounding sediments, scales, fins, and soft bones retain their positions and provide the necessary resistance to leave an impression of the body form when the flesh is gone.

Insect Fossils (enlarged)
Fine specimens of this type are obtained from an old lake bed at Florissant, Colorado.
The larger and more spectacular fossils, such as skeletons, skulls, and detached bones are nearly always of the replacement type. Replacement of plant and animal substances by mineral matter is a slow process and in younger fossils the change is rarely completed, some of the original material being present in a partially altered condition or not modified at all. Since air does not often carry the necessary materials and provide other essential conditions, replacement may be regarded as something which happens underground or in water. It is perhaps best explained in connection with limestones, because calcite or “lime” is frequently the replacing substance although other minerals, especially quartz, may serve the purpose.
Besides converting bony or woody objects into rock substance, mineral replacements may assist in the production and preservation of fossils in another manner. It often results in the filing of cavities with some rock-making substance which retards destruction through crushing or other injury. In many cases, so-called fossil shells are not shells at all; instead, they are merely a stony filling which was once surrounded by 11 shell substance. In other instances the original shell remains as it was during the life of its former occupant, preservation of the shell being due largely to the substitution of a mineral filler for the soft animal tissues once present.
Limestone comes into existence through a more elaborate process than that which produces sandstone and shales. It is one of the three types of common rocks, known collectively as the sedimentaries, in which fossils are found. It differs from sandstones and shales, however, in that much of its substance has been dissolved in water instead of being transported in the form of finely ground rock particles. Lime occurs in many varieties of rock which are exposed to the wear and tear of the elements throughout the world. Slowly but more or less continuously it is taken from this source by ground and surface waters coming in contact with it. Particularly active is carbonated water, moving underground through pores and crevices.
This underground circulation of mineral matter in a dissolved condition explains the occurrence of fossils in land areas which have not necessarily been submerged during any great length of time, for it is well known that plant and animal remains are not invariably washed into lakes or seas, and that all sedimentary deposits have not been built up in large bodies of water. Here we are dealing with what is known as the continental type of sedimentation and such fossils as dinosaurs, mastodons, three-toed horses, and other former inhabitants of land areas.
In order to become properly fossilized, certain conditions are absolutely necessary, and only a small percentage of the once-living multitude secures the required treatment. There must be present, soon after death, some protection from the activities of the carnivorous birds and beasts that would separate and scatter the parts of a carcass, also from the smaller gnawing animals that would continue the destruction, and finally from wind, sun, rain, frost, and bacterial and chemical activities which in the course of only a few years would remove everything but possibly a few scraps of tooth enamel, which is the hardest of animal tissues.
A slight covering of earth substance in any form serves to check the disintegration, and this may be acquired in several ways. Animals that perish in bogs or quicksands are soon covered over; in many localities wind-blown dust and sand do the work; and flooded river valleys provide an abundance of mud for the necessary burial of others. Even underground, the decay of soft tissues is too rapid to permit of replacement by mineral substance in a manner that would reproduce form and texture. Skin and flesh are almost invariably lost, although in a few instances the thick scaly hides of dinosaurs are known to have produced natural molds and casts by the method explained in connection with footprints and other impressions.

The skeleton of this prehistoric American rhinoceros is mounted in a “half shell” which was modeled over the bones to show the form of the living animal. The artist’s reconstruction appears in the painting above the fossil exhibit.
With regard to the more durable tissues found in the teeth, bones, and shells of animals, or the woody parts of plants, the case is different. These parts become firmly imbedded in the ground, but moisture still has access, and it begins to work immediately; for all water moving underground finds soluble substances which it picks up and carries with it wherever it goes, and much of the load consists of mineral matter which may be unloaded again when the necessary conditions are found.
Mineral-laden waters will drop one kind of substance to take up another which dissolves more readily, and this happens sooner or later when a buried bone or log is encountered. Complications of various sorts enter into the process, but the final outcome frequently is a complete change from one chemical composition to another which is more enduring, the transformation being brought about so gradually and thoroughly that in many fossils the inner structure of the original tissue is as accurately reproduced as the fine detail of surface features.
Converted into stone, however, the result is still far from permanent. While yet underground the fossil is subjected to distortion and breakage due to earth movements which bend and dislocate the rock deposits. What causes these upheavals and depressions of the earth’s surface remains the subject of much discussion, but that they have occurred on a large scale and continue to occur is clearly evident. At higher altitudes the surface rocks and fossils are exposed to a larger variety of destructive activities than at lower levels where protective coverings are more likely to be provided and retained. Once stripped of that protection there is little chance for a fossil to survive. Beyond a doubt there are many thousands of tons of prehistoric remains damaged or destroyed each year, by weather and stream erosion.
As the various types of sediments continue to accumulate on land and in water they produce deposits of sandstones, claystones, and limestones which in time may acquire great thickness and cover wide areas of sea floor, or continental surface. Usually there is more or less mixing of sediments resulting in sandy limestones, limy clays, and other combinations. Quite commonly, however, the types remain fairly pure but become arranged in layers which alternate from one kind of material to another. At all times the character of the deposit will depend upon the nature of the rocks which supply the materials, and any fossils that may be produced will consist of such plants and animals as live and die during the time the rock is in the making.
Some of the rock layers will be rich in plant and animal remains, others quite barren, the difference being due partly to conditions influencing the life of the region. In addition, the character and amount of rock-making materials at the time may be favorable or unfavorable to the preservation of fossils. Seas, lakes, and valleys may at any time be drained, or enlarged and deepened, by changes in the elevation of underlying rocks. The amount and variety of mineral substances dissolved in the waters of a region not only affect the character of rock deposits but also the plants and animals living in the water. Some of these chemical solutions provide cementing materials which bind together the grains of sands and mud; others have a detrimental effect upon cementing material previously deposited, and so construction and destruction go on continuously, more or less hand in hand, to produce complicated and often puzzling results.
A little more salt, or a little less of it, may change completely the variety of life inhabiting a body of water. A slight change in the depth of the water often accomplishes the same thing, for plants and animals are so delicately adjusted to their environments that conditions fatal to one race of creatures may provide the exact life requirement of another. This is a matter of practical knowledge which is being used today in the cultivation of plants and animals for market purposes. It is being demonstrated continuously, also, upon living subjects in experimental laboratories throughout the world; and, in a bigger way, the facts are observable wherever life is considered in relation to habitat. That anything so obvious should be regarded as guesswork or theorizing, or opposed to truth, when applied to former inhabitants of the earth, is somewhat surprising. And, it may be added, the cultural worth of fossil study comes to a focus on this very point, for men and women are now meddling, consciously or unconsciously, wisely or unwisely, with an all-important environment about which they have learned very little—one called, among other things, “civilization.”
For any portion of the world a complete-list of the different kinds of plant inhabitants comprises the flora of that region, and a like summary for the animal life is known as the fauna of the district. It is generally understood that different species of both plants and animals inhabit different regions of the earth, but outside of professional circles it is only beginning to be recognized that changes in floras and faunas occur from time to time, that slight differences may be noted in the course of observations extending over a period of only a few years, and that everything in a fauna or flora eventually may be displaced by new forms.
It is, however, a convenient practice to use these terms in connection with time periods, rock beds, and types of environment, as well as geographical areas. Thus we have such phrases as a “Cretaceous fauna” (attaching the name of a geologic period), a “Benton fauna” (with reference to the fossils of a rock formation), a “marine flora” (using the name of an environment), an “Arctic flora” (which applies to a definite portion of the earth surface and its plant inhabitants).
Faunas include animals which many persons do not recognize as such. Sponges, corals, insects, worms, crabs, oysters, and a host of other boneless creatures are grouped together as invertebrate animals, while another group includes the fishes, amphibians (toads, frogs, and salamanders of today), reptiles (crocodiles, lizards, snakes, and turtles being well known varieties), birds, and mammals. This second lot, provided with backbones and skeletons, comprise the great division of vertebrate animals.
Floras also include types which are commonly seen but not popularly identified as plants. The algae are perhaps best known as seaweeds, water-silk, and pond scums; fungi as toadstools and moulds. Both groups are large and of important rank in the vegetable kingdom; only the algae, however, are recognized as important fossil producers. Better known types of plants are the mosses, ferns, evergreens, grasses, and the more conspicuous flower-bearing forms, from weed size to tree size.
Many rocks owe their character to the work of large colonies of plants or animals, for the living organisms are frequently the active agency which takes dissolved mineral substance from the solvent liquid and gets it back into solid form. The liquid is, of course, the water in which the creatures live, while the mineral substance often becomes a commodity required by a plant or animal in its mode of living. Mollusks have a way of using lime in the production of shells, and many a bed of limestone consists almost entirely of this by-product of molluscan life. Tiny coral polyps build complicated and beautiful structures from the same mineral substance. Either intact or in broken condition, these structures contribute in a large way to the making of limestones. Algae, among the lowliest of plants, have done extensive work along similar lines, and numerous invertebrate animals could be named as important factors in the production of rocks. Many of the shells and other fabrications retain their peculiar patterns long after the extermination of their makers, and a highly informative part of the fossil record is provided in this manner. It is also by far the larger portion of the record, for the earlier ages of prehistoric time failed to produce a vertebrate animal of any kind, while the invertebrate record dates back to pre-Cambrian time.
If in some part of North America there had been steady accumulation of sedimentary materials under constantly favorable conditions since the beginning of Cambrian time, the result would have been a deposit of sandstones, claystones, and limestones measuring nearly fifty miles from bottom to top. These figures are based on actual production in North America where extensive measurements have been made in many localities. When other parts of the world are as thoroughly investigated and older deposits included in the calculations, the total thickness of such beds will probably be more than one hundred miles.
No single pile of rocks offering a complete cross section of the geological record has ever been produced, but portions of the section are exposed to view on all the continents. In order to carry on desirable investigations and make comparisons, it has been necessary to divide this great composite section into small units which may be named in some way and placed definitely with relation to lower and higher, or older and younger, layers. To serve this purpose there has been developed the idea of rock formations, and here we have a word which is not defined readily, even for the use of those who are familiar with it. Nevertheless it is used so commonly that some understanding of its meaning becomes desirable.
A formation may be regarded as an extensive rock mass, variable, in thickness and other proportions, as well as in composition, but representing a period of time during which there was no great change in the character of plant and animal life, and no serious interruption in the depositing of the rock-making materials. Occasionally the lower and upper limits of a formation are well defined and readily located. Frequently, however, the transition is gradual, one formation merging into another with no apparent mark of separation. In such event the original description serves to establish more or less definitely the boundaries of a formation.
Descriptions are published whenever a worker believes he has discovered a significant part of the great section which has not previously been named. The usual practice is to apply a name taken from the locality in which the beds were investigated, and in this manner the names of formations become associated with towns, rivers, counties, mountains, states and other geographical features. The locality which supplies the name is then regarded as the “type locality” for the formation, but wherever these same beds may be traced or otherwise identified the one formation name applies.

Dinosaur Tracks
An ancient trail in sandstone of the Dakota formation. East slope of the hogback, west of Denver.
The “Dakota formation,” to use a convenient illustration, is mentioned in scores of reports bearing on the geology of Colorado, Iowa, Kansas, Nebraska, New Mexico, Texas, Utah, and Wyoming, as well as the Dakotas. On the geological map of Colorado it appears on both sides of the Rockies, scattered in strips and patches from north to south boundary lines. The beds are easily located in the foothills district west of Denver because of their tendency to produce the prominent ridges known as hogbacks.
Many formations are exposed over much less territory, some have even greater extent. Thickness may vary from a few inches to thousands of feet, and no two exposures will be exactly alike though some similarity necessarily prevails throughout. “Exposures” are simply portions of the beds which are not concealed by loose rock, soil and vegetation, or overlying formations. Canyon walls, steep cliffs and mountain slopes, gullies, and badlands provide a large variety of natural exposures. In such places rocks and fossils may be studied to best advantage.
Since a formation may contain a variety of beds, including sandstones, shales, limestones, and all sorts of mixtures, there is sometimes need of subdividing it; but formations are the smallest units commonly shown on geological maps. They are actual rocks which fit into a historical scheme of things and may be regarded aptly as the pages of a book which nature has done in stone.
“How old are they?” “How can you learn their names from the rocks?” These are typical examples of questions most frequently asked concerning fossils. The second question follows the usual reply to the first, for prehistoric plants and animals are as old as the rocks in which they are found. The answer, as to age, must come from the rocks and what we have learned about them through many years of hard work, thoughtful observation, and careful study. Names, however, come from a different source. Nature, apparently, managed for a long time to carry on without the use of words. Since man began talking he has had no trouble inventing names for things which interest him.
Early students of rocks and fossils likewise accomplished a great deal without being able to date events in terms of years although many of their efforts and interests centered on the problem of discovering a continuous sequence of events in the fragments of evidence that had been uncovered. This relatively simple problem has not been fully worked out, and some of the breaks in the record are recognized as “time gaps” which may never be converted into history.
The question of time, expressed in years, has been a puzzle which attracted some attention even in the earliest days of investigation. Its solution was attempted by several methods long before there was sufficient information to make them work satisfactorily, which accounts in part for the extreme variation in results of the calculations. Even now it is to be expected that changes will have to be made as long as pertinent studies are continued. Two of the most promising methods of investigation in late years have been producing figures which are surprisingly large. More accuracy than ever before is probably present in modern estimates but, except for comparatively recent time, there is yet no way of knowing within a range of millions of years when a creature lived.
Astronomy and physics were used in early calculations but, although taken seriously by some geologists, it was soon recognized by others that certain events revealed by earth history could not be explained with so short a time allowance as these methods indicated. One of the first estimates provided a total of only twenty-five millions of years and included a great stretch of time during which the earth, according to prevailing theory, was more sun-like than rock-like, a time when planets were being born and the earth could not have been in its present physical condition, which is the chief concern of the geologist. Since those earlier conditions could not have supported life as we know it, our knowledge of cosmic history renders small service in the study of fossils.
Among the methods suggested by astronomy and the laws of physics is one which is based on the probable rate at which the earth cooled from its molten condition to present temperature. It is believed now that the heat of the earth is not necessarily due to an original molten state and that a steady rate of cooling cannot be ascertained. Any figures based on such procedure, therefore, are discredited today.
The amount of salt in the oceans, and the time required for its concentration there by natural processes, offers another way of attacking the problem. It is a well known fact that salt is being added to the seas at a fairly constant rate; sea water, then, must become saltier from year to year. The salt comes from rocks exposed on land surfaces and is transported by the rivers which drain these areas. By analyzing the river waters it is possible to estimate the amount of salt annually dumped into the oceans and, also by chemical analysis, it is a comparatively simple matter to figure the total amount now present in the oceans. Some recent calculations indicate that thirty-five million tons of salt are being added each year, and this figure divided into the total amount for all the years places the age of the oceans at three hundred sixty millions of years.
However, there are certain other factors which complicate the problem. For instance, it is known that land areas exposed to surface drainage 20 have not always been of their present size, and the annual production of salt by the different types of rocks exposed at various times in the history of the earth has not always been as it is now. The rocks also must be older than the oceans, but how much older cannot be determined by means of figures obtained in this way.
Until the beginning of this century there was little anticipation of a better measuring stick than one in use at the time which placed its reliance on the total thickness of the sedimentary deposits and the length of time required to produce this great accumulation of material which is known as the geological column. Since the total thickness, or height of the column, was not accurately known, and with recognized time gaps to bridge, there was little hope of working out a complete chronology by this device, but it has supplied highly desirable and reliable information concerning parts of the record.
The system has been somewhat improved since its earliest use, and one of its latest applications gives us an age, for known sedimentary rocks, of nearly half a billion years, this being based on a total thickness of one hundred miles and an average rate of 880 years for the building up of one foot of sediments. Its greatest weakness is due to the absence of a reliable factor to take care of long stretches of time in which the sedimentary rocks are known to have been subjected to destructive processes. A yardstick of this character cannot be applied to rocks that have been destroyed, and there are excellent reasons for believing that these interruptions may account for several times the lapse of years indicated by the amount of rock remaining in the column which has been pieced together.
Following the discovery of radium, however, the present century provided a new field of knowledge which has contributed greatly to the measurement of geologic time. The penetrating rays produced by radium and other radioactive substances are due to extremely slow but violent disintegration of the material. Uranium and thorium are radioactive elements which occur in the rocks of many parts of the world. There is little or no loss of material as the so-called disintegration proceeds; instead there is a complicated series of transformations in which other elements are produced, radium itself being one of these. Helium and lead eventually take the place of the less stable elements and the known rate at which these products accumulate provides the highly desired key to the age of the rocks.
Part of the gas, helium, may escape, but except in rare instances where chemical alteration might occur, there probably is no loss of lead. Fortunately, when this metal is produced by radioactivity it differs slightly in atomic weight from ordinary lead; otherwise the presence of the latter 21 would introduce a misleading factor. Since the speed at which the change goes on cannot be increased or decreased, it is assumed that throughout past ages it has never been faster or slower. The amount of such change that has been completed in any body of radioactive minerals may be measured by techniques employed in physics and chemistry. If it is found that the amount of helium or lead present requires a hundred million years for its production at the working speed of the parent elements, the mineral deposit must be at least that old.
Certain conditions of course complicate the problem seriously: knowing the age of a piece of rock which happens to contain some radioactive element is of small service in historical studies unless the rock can be definitely associated with a flora or fauna, or some outstanding event disclosed by geological investigations. But there have been a few instances in which most of the necessary conditions were present, and more and better opportunities to apply this method will no doubt appear. Other elements, or their radioactive isotopes, are already being employed with good results. Some of these, such as carbon 14, are more sensitive indicators for the accurate dating of events in comparatively recent time.
When it can be used, this type of measurement is far less subject to uncertainties than any other. It promises to eliminate all need for guessing, and comes close to a degree of accuracy which is satisfactory to the scientist, a person who thoroughly dislikes uncertainties of any kind. If suitable material can be found in just the right places it should accomplish what the preceding method cannot do—the accurate measurement of the great time breaks which interrupt the geological record in many places. Something along this line already has been accomplished, for radioactive material has been found in some of the oldest of the rocks. Regardless of the destruction going on in other localities, these rocks have continued to register the passing of time, and a tremendous antiquity for the earth and some of its first inhabitants has been indicated.
Tests made on radioactive minerals from Gilpin County, Colorado, have established the age of late Cretaceous or early Cenozoic rocks at sixty million years, providing a convenient and reasonably accurate date for the beginning of the Age of Mammals. In Russia, one of the oldest mineral deposits yet studied in this way and regarded as early Pre-Cambrian, produced the astonishing figure of 1,850,000,000 years; what we commonly refer to as geological history may therefore be regarded as covering a range of approximately two billions of years. The earth, in some form or other, has in all probability passed through an earlier history of another billion years or more.
Wherever we may roam, a portion of the prehistoric record is to be found in the rocks underfoot and not far from the surface. Formations as already mentioned may be regarded as the pages—often torn and badly scattered—of nature’s own book, in which the geological periods are chapters. But instead of numbering these pages and chapters we have named them, in order to get the parts reassembled in orderly fashion and restored to a condition which makes the book legible. However, the names cannot render the service intended except in connection with a time chart and an outline of earth history.
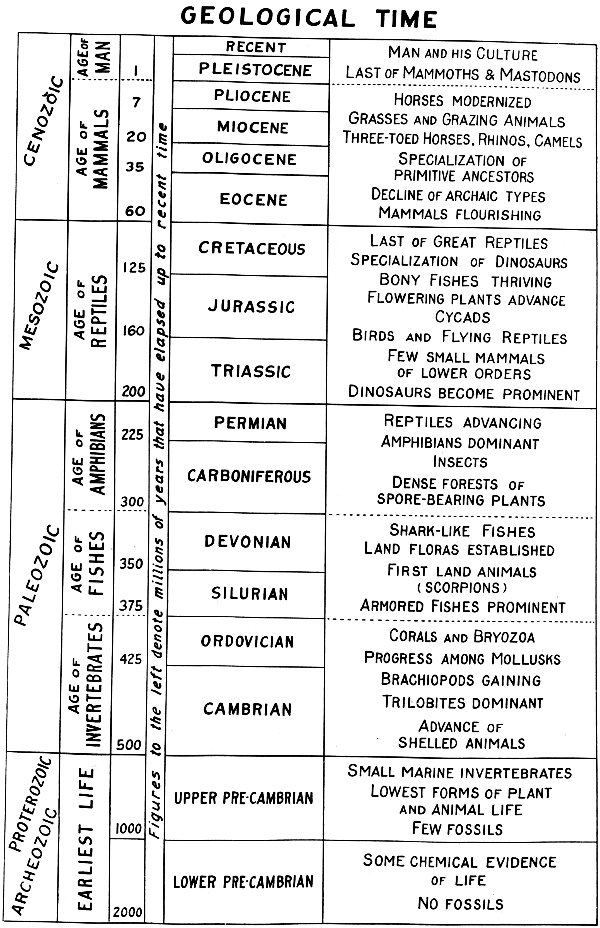
GEOLOGICAL TIME
Figures to the left denote millions of years that have elapsed up to recent time
| CENOZOIC | ||
| Age of Man | ||
| RECENT | Man and his Culture | |
| 1 | PLEISTOCENE | Last of Mammoths & Mastodons |
| Age of Mammals | ||
| 7 | PLIOCENE | Horses modernized |
| 20 | MIOCENE | Grasses and Grazing Animals |
| Three-toed Horses, Rhinos, Camels | ||
| 35 | OLIGOCENE | Specialization of Primitive Ancestors |
| 60 | EOCENE | Decline of archaic types |
| Mammals flourishing | ||
| MESOZOIC | ||
| Age of Reptiles | ||
| 125 | CRETACEOUS | Last of Great Reptiles |
| Specialization of Dinosaurs | ||
| 160 | JURASSIC | Bony Fishes thriving |
| Flowering plants advance | ||
| Cycads | ||
| Birds and Flying Reptiles | ||
| 200 | TRIASSIC | Few small mammals of lower orders |
| Dinosaurs become prominent | ||
| PALEOZOIC | ||
| Age of Amphibians | ||
| 225 | PERMIAN | Reptiles advancing |
| Amphibians dominant insects | ||
| 300 | CARBONIFEROUS | Dense forests of spore-bearing plants |
| Age of Fishes | ||
| 350 | DEVONIAN | Shark-like Fishes |
| Land floras established | ||
| 375 | SILURIAN | First land animals (scorpions) |
| Armored Fishes prominent | ||
| Age of Invertebrates | ||
| 425 | ORDOVICIAN | Corals and Bryozoa |
| Progress among Mollusks | ||
| 500 | CAMBRIAN | Brachiopods gaining |
| Trilobites dominant | ||
| Advance of shelled animals | ||
| PROTEROZOIC | ||
| EARLIEST LIFE | ||
| 1000 | UPPER PRE-CAMBRIAN | Small marine invertebrates |
| Lowest Forms of Plant and Animal Life | ||
| Few Fossils | ||
| ARCHEOZOIC | ||
| 2000 | LOWER PRE-CAMBRIAN | Some chemical evidence of life |
| No fossils | ||
Such aids have been devised and revised from time to time. No figures have been offered as final or absolutely “right” since the beginning of scientific investigations. Time divisions have been proposed that are not yet in common use while others have been abandoned or modified. Sources of information are so numerous that appropriate credit cannot be given fairly for anything that is up-to-date. The combined chart and outline here provided is based on time calculations of recent date but with figures slightly rounded off for the sole purpose of making them easier to remember. In view of the still existent probability of error it is felt that the slight alteration of figures may justify itself. It need not be regarded as misleading if the present purpose be considered—the stimulation of a natural history interest which is not vitally concerned with the little difference between a thousand million years and nine hundred ninety-nine million years.
The whole of geological time has been divided and subdivided according to varying practices. The development of life is perhaps the one outstanding feature of the time divisions, but for the most part the changes in floras and faunas have been gradual rather than abrupt, and this makes it very difficult to draw sharp lines or to visualize beginnings and endings of the various stages of development. Occasionally there is good excuse for drawing a line, where the record is broken and resumed again after a long lapse of time. The principal cause of such breaks is the elevation of great land masses, which brings on an interval of erosion and surface destruction for the areas uplifted.
These movements of parts of the earth’s crust have been exceptionally pronounced at certain times, often culminating in the production of mountain systems, and because of the extreme changes they introduce are known as revolutions. The major divisions of prehistoric time have been established, at least in part, by such revolutions; crustal, climatic, or other disturbances, on a smaller scale and recurring with greater frequency, may be regarded as establishing boundaries for the minor divisions. Hence we have five great Eras of geological history, and 24 these are divided again into Periods. The time chart shows an arrangement commonly used in America. In the first column the names of the Eras are stated in technical form. Closely coinciding with these terms are the popular names of the Ages which appear in the second column. These names, describing the dominant life of each age, are very convenient. The more scientific terms used for the eras, while serving essentially the same purpose, are a little more systematic and generalized in that they refer to ancient life (Paleozoic), middle life (Mesozoic), and recent life (Cenozoic), without being specific as to any class of animals or plants for any one division of time.
The period names, in the central column, have been derived from miscellaneous sources, some of them from geographical districts, some from descriptive references to prominent features of the rocks, others indicating a degree of approach to recent time. In paleontology (fossil study) it has long been a practice to cut the periods into lower, middle, and upper divisions, and in a few cases it has been found desirable to make two periods out of an old one. What was once known as the Lower Carboniferous is now commonly recognized as the Mississippian period while the upper portion has become the Pennsylvanian. The Lower Cretaceous is now the Comanchean of some authors.
Both old and new practices are responsible for a little confusion at the present time. A former division into Primary, Secondary, Tertiary, and Quaternary eras has been partly abandoned, but the term “Quaternary” still applies to the Age of Man, while “Tertiary time” remains in good usage for the balance of the Cenozoic era. Among the newer introductions may be mentioned the use of a Paleocene period which precedes the Eocene. Geologists are not entirely in agreement as to the necessity for this addition and many would not give it equivalent rank with other periods. In the interest of simplicity these modern refinements have been omitted from the chart.
The figures appearing in the third column, between the Ages and Periods, indicate the millions of years that have elapsed up to present time. They denote the age of the rocks at the beginning of each period. The age of a plant or animal which lived in Eocene time would be, according to this scale, somewhere between 35 million and 60 million years. In practice it is usually possible to determine whether a fossil was embedded in the rocks during an early or late portion of the period, and thus its age may be established within a shorter range, but it is impossible to be exact, even in terms of millions of years, with regard to anything as far back in prehistory as the Eocene period.
The period in which we are living today is known as Recent. It began at the close of the Ice Age or Pleistocene period about ten thousand 25 years ago and represents so little of earth history since the beginning of life that a chart many times the length of this page would be required to show the rest of the periods in proportion. The Cambrian period is an early chapter in which the story of prehistoric life suddenly becomes clear and richly varied. It is, however, much farther from the beginning of the record than it is from the present, and the Pre-Cambrian eras would require a great deal more space in order to show their relative lengths. The Archeozoic and Proterozoic eras have to some extent been divided into periods, but the great antiquity of the rocks has obscured much of their history, and divisions established for one locality have been of little service elsewhere. Consequently, the period names are in less general use and the common practice is to refer to all this great stretch of time as Pre-Cambrian.
In the last column, at the right of the chart, some of the historical features are indicated. This column should be read from bottom to top in order to get the proper development of the story, and at best this sketchy outline of events can be no more than suggestive of the progress and decline through which the earth’s inhabitants have passed.
Rocks of every period except probably the Silurian are known to have been deposited somewhere in the Colorado area, although in most cases the record for each period is far from complete. Formations are too numerous and too varied locally to be shown on a chart of this type.
In the study of fossils there are two important field aids usually available. For any locality there should be a geological map and a section showing the sequence and character of the strata. On a small-scale map many of the local details have to be omitted, but the position of the larger exposures is indicated and, with this information at hand, the fossil-bearing strata may be located with the help of a geological section. The latter is frequently obtained from technical reports published by State and National Geological Surveys. Frequently, however, it is possible to obtain only a general plan for a given locality, and a great deal of literature may have to be scanned in order to get that. Excellent geological maps of Colorado have been published by the Colorado Geological Survey and the United States Geological Survey.
It often happens that a formation is not where we expect to find it, this being due to several possible factors. The sediments may not have been deposited there, or they may have been removed by erosion. Where the structure has been disturbed by folding and faulting, a multitude of complications is introduced. The expected sequence is sometimes inverted and repeated through a series of folds. Formations also may be moved miles out of place by faulting. Both thickness and character of sediments may vary considerably within a formation. In some regions the geology is very simple, in others extremely difficult to understand.
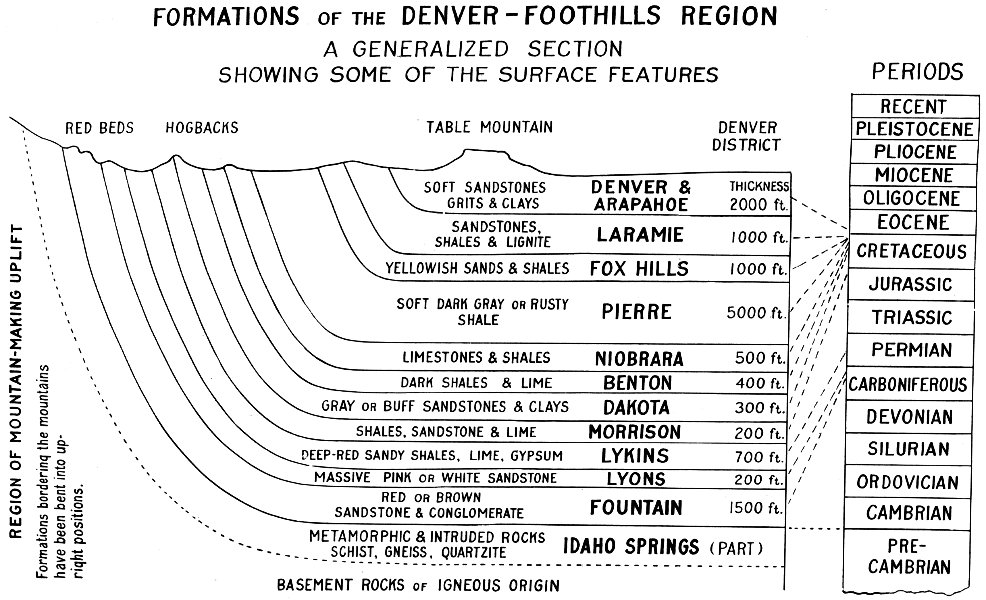
FORMATIONS of the DENVER-FOOTHILLS REGION
A GENERALIZED SECTION SHOWING SOME OF THE SURFACE FEATURES
RED BEDS HOGBACKS TABLE MOUNTAIN DENVER DISTRICT
REGION OF MOUNTAIN-MAKING UPLIFT
Formations bordering the mountains have been bent into upright positions.
| PERIODS | ||||
|---|---|---|---|---|
| RECENT | ||||
| PLEISTOCENE | ||||
| PLIOCENE | ||||
| MIOCENE | ||||
| OLIGOCENE | ||||
| EOCENE | ||||
| CRETACEOUS | THICKNESS | |||
| SOFT SANDSTONES GRITS & CLAYS | DENVER & ARAPAHOE | 2000 ft. | ||
| SANDSTONES, SHALES & LIGNITE | LARAMIE | 1000 ft. | ||
| YELLOWISH SANDS & SHALES | FOX HILLS | 1000 ft. | ||
| SOFT DARK GRAY OR RUSTY SHALE | PIERRE | 5000 ft. | ||
| LIMESTONES & SHALES | NIOBRARA | 500 ft. | ||
| DARK SHALES & LIME | BENTON | 400 ft. | ||
| GRAY OR BUFF SANDSTONES & CLAYS | DAKOTA | 300 ft. | ||
| SHALES, SANDSTONE & LIME | MORRISON | 200 ft. | ||
| JURASSIC | ||||
| TRIASSIC | ||||
| PERMIAN | ||||
| DEEP-RED SANDY SHALES, LIME, GYPSUM | LYKINS | 700 ft. | ||
| CARBONIFEROUS | ||||
| MASSIVE PINK OR WHITE SANDSTONE | LYONS | 200 ft. | ||
| RED OR BROWN SANDSTONE & CONGLOMERATE | FOUNTAIN | 1500 ft. | ||
| DEVONIAN | ||||
| SILURIAN | ||||
| ORDOVICIAN | ||||
| CAMBRIAN | ||||
| PRE-CAMBRIAN | ||||
| METAMORPHIC & INTRUDED ROCKS SCHIST, GNEISS, QUARTZITE | IDAHO SPRINGS (PART) | |||
| BASEMENT ROCKS of IGNEOUS ORIGIN |
A generalized section for the western part of the Denver Basin is introduced here for the use of local students. The formations normally present in this region are shown in their usual position. They are briefly described on the chart, and their thickness is indicated by figures which may be regarded as near the maximum for the district. The section will apply to most of the foothills area between Morrison and Boulder though surface features and thickness of beds will vary considerably from place to place.
Certain of the formations are known to be fossil bearing, others barren or nearly so. When fossils are present they are usually restricted to certain localities, and these may be widely scattered. The following remarks apply to the possibilities for finding fossils in the formations named.
Leaf impressions of palms, ferns, and numerous species of well-known trees and shrubs are common in many localities. Petrified wood is fairly abundant, and a few scattered bones of reptiles and mammals have been found. The two formations are treated as a unit because the Arapahoe is neither conspicuous nor sharply defined. Denver beds are well exposed on the slopes of Table Mountain at Golden; fossils, however, have been obtained from several localities nearer the city of Denver, notably from the hills just west of Overland Park.
Plant material is locally abundant, principally the leaves of familiar deciduous trees, palms, and ferns. Many of the clay pits being worked near Golden are in this formation. Oysters and a few other mollusks may be found in some places.
Better exposures of this formation are located to the north of Denver. Marine mollusks are most frequently found.
In addition to the characteristic dark shales, this formation includes some limy material and sandstone beds, both of which are fossiliferous in places. Two types of marine mollusks are characteristic: Inoceramus, generic name for several species of clam-like bivalves readily identified by concentric elevations which produce a rippled effect on the shell surfaces; and Baculites, cephalopods with straight, chambered shells which often break at the suture lines, where the fossil is weakened by the chamber walls. Small oyster shells are fairly common also. The formation is to be found some distance to the east of the prominent hogback where it weathers into smooth surfaces in the form of broad valleys and flats, with rounded contours on the few elevations that may be present. It forms a soft, flaky soil when dry, is a sticky “gumbo” when wet. The clay is generally of a rather dark grayish color when freshly exposed but it takes on a rusty appearance after weathering. At various levels there are numerous iron-cemented concretions, many of which contain fossil shells.

Marine Beds of the Benton Formation, Northeastern Colorado
A stream channel has cut deeply into the formation, uncovering and partly destroying a plesiosaur skeleton which was found at the level where the men are standing.

Plesiosaur Bones in Place
Benton formation. Surface rubble has been cleared away, and several vertebrae are partially uncovered in the area at the right of the hammer.
The formation contains fossils rather similar to those of the Pierre. Shark’s teeth have been found in some of the lower beds. Limestone is a prominent feature, often forming a well defined ridge near the foot of the eastern slope of the main hogback. The limestones commonly have a chalky character.
The formation is not especially productive in this region. Marine shells are numerous in some localities, and bones of marine reptiles have been found at various places. As usually seen, it is almost entirely composed of impure clay shales, very dark, brownish-gray to almost black, and commonly interbedded with thin patches of white bentonite, yellow ochre, gypsum, and limestone.
This formation produces the high hogback which is usually present some distance east of the Red Rocks. There are generally two or three layers of massive, light-colored sandstone separated by clays which are used extensively in the making of bricks and pottery. Leaf impressions and some fish scales are found in the clays and occasionally in the sandstone. The hogback is a good marker from which to locate other formations, because of its prominence in the foothills landscape.
Good dinosaur material has been taken from the Canon City and Morrison districts. The formation is to be found on the lower west slope of the Dakota hogback. It consists of continental deposits of the stream and lake types. There is considerable sandstone in this formation and a little limestone is to be found here and there, but the most characteristic feature is in the shales. When freshly exposed, the shales are delicately tinted with gray, green, and maroon, a bronze-green being rather prominent. This formation is highly variable in character, with much of the clay often buried under the valley floor. In addition to the bones of reptiles, there are plant fossils, usually of poor quality, and fresh-water gastropods more or less abundant in some localities.
Outcrops are not prominent, owing to the small amount of weather-resisting materials. The sandy clays are commonly of a deep red 30 color mottled with spots of light gray. A white limestone is sometimes present near the middle of these deposits, and gypsum beds are included locally. The formation is often indicated only by red soil in the depressions between ridges. Few fossils have been reported.
This formation is usually prominent as the eastern wall of the uplifted Red Rocks series. In some localities it forms a ridge of pink or white sandstone distinctly separated from the older sediments to the west. Very few fossils are found.
Exposures usually are brown to red in color, though sometimes a dirty white. The prominent rocks are rather coarse sandstone, commonly with a gritty texture due to the angular character of the sand or gravel from which they were made. These are the westernmost of the Red Beds and the oldest of the uplifted sedimentary rocks bordering the foothills in most of our area. Fossils have been found in the formation, but it is practically barren for the territory here considered.
* * * * * * * *
This geological section also illustrates a method of dating crustal movements and the birth of mountain ranges, for the folding of the strata along the flanks of the Rocky Mountains has a great deal of significance in this connection. The sedimentary layers were originally deposited over much of the present mountain area in a horizontal position, and only those formations in existence at the time could be distorted by the upheavals which produced the new elevations. Of the series generally involved in the movement the Laramie beds are the youngest. Since these beds had not been formed until near the close of the Cretaceous period it is to be assumed that the mountains must be of more recent date, younger than the topmost of the deformed beds and at least as old as the lowermost of the undisturbed formations overlying them.
Some disturbance is evident also in the Arapahoe and Denver beds which overlie the Laramie, but this is believed to have occurred sometime after the occasion of the first great uplift. Volcanic materials in these beds lead to the belief that the sediments were deposited during a period of volcanic activity brought on by the crustal folding which terminated the Mesozoic era. Hence the conclusion arises that the age of the Denver and Arapahoe beds must coincide closely with some of the earlier stages in the history of the mountain system. This interval is often referred to as Post-Laramie time.
The rocks of Pre-Cambrian time have been buried deeply under the accumulation of younger sediments, and the resulting pressure in many places has been tremendous. In addition to the effects of pressure there also is recorded in these ancient formations the repeated movements of the materials since they were first deposited. Vertical and side adjustments of parts, with relation to other parts, have distorted the original arrangement of the rock particles to such an extent that ordinary fossils would eventually become unrecognizable. These crushing, grinding, and kneading forces working through millions of years alone would account for the absence of fossils from the older deposits. Frequently the rocks have become so changed in form that their original character can only be conjectured, and because of this change they are known as metamorphic rocks.
A few beds of Archeozoic age remain in nearly their original condition, but they are either without fossils or they have produced very questionable and unsatisfactory specimens. The existence of life during these early stages of earth history is indicated largely by chemical rather than fossil evidence. Much of the ancient limestone has been converted into marble, but it is not unreasonable to believe that plants and animals were instrumental in the production of this type of rock as they are today. Certain varieties of iron ore deposits are now being built up by the aid of plants, and similar ores in the ancient rocks may have had a like origin. The presence of great quantities of carbon, in the form of graphite, may be regarded also as a sign of life, for this substance is accumulated on a large scale by living plants, and may be retained in a solid form after the partial decay of the plant tissues.
So far as the direct evidence goes, there is no sign of any creature of large size or of such complicated structure as the common plants and animals of today. The chemistry of the mineral deposits is not entirely convincing as to the presence of life, but it is regarded as highly probable that microscopic, single-celled plants and animals, comparable to modern algae and protozoa, were in existence during Archean time. Throughout later eras there is unmistakable evidence of gradual development from simpler to more elaborate life-forms and the Archeozoic is commonly regarded as a time of preparation during which simple organisms of some kind were becoming adapted to early conditions which could 32 not support life on a higher plane. The importance of the work done by such lowly creatures in the preparation of suitable environments for more advanced modes of living is overlooked almost entirely.
During the next era, the Proterozoic, the record of life becomes somewhat clearer. Fossils are hardly to be regarded as abundant but there were several well-defined types of animals which left shells and other parts composed of mineral matter. Among these may be mentioned the Radiolaria, Foraminifera, Bryozoa, and Sponges. Radiolaria produced delicate, often lace-like shells of many patterns adorned with the radiating filaments or spines which have suggested the name for this group. Foraminifera produced minute shells, sometimes many chambered, and often bearing a confusing resemblance to the work of snails. Common chalk is composed almost entirely of such shells and fragments of them.
Sponges and Bryozoa are animals of slightly higher organization. They are many-celled instead of one-celled and the cells have special work to perform, which is a most important step in the direction of the specialization which characterizes the structural and life pattern of later arrivals. The Bryozoa lived in moss-like colonies which have been important rock-makers; the fossil forms bear some resemblance to corals. Sponges are too well known to require description although the familiar article of commerce is merely the framework of once-living animals. They represent the earliest organization of true animal bodies even though in appearance they may have a resemblance to plants.
Actual plants of this era were of the algae class, aquatic in habit as were their animal neighbors, the first to leave a record in the form of fossils. This record, obscure and distorted, has long been a source of perplexity to investigators. Without well-defined floras and faunas to guide them, and with rocks frequently in chaotic relationships, early geologists were content to regard it all as a “Pre-Cambrian complex.” Recent studies have contributed a great deal of information not available some years ago. It is quite possible that more advanced types of life were in abundance before the close of the second era, but material on which to base sound opinion is still scarce.
Rocks of Pre-Cambrian age are plentiful in the foothills region west of Denver. The schists, gneisses, and quartzites exposed for some miles immediately beyond the red-beds are part of this great complex. The Idaho Springs formation is known to be one of the oldest in this district, although its exact age has not been determined. Other formations are recognized among the metamorphic rocks of the region but none has contributed to our knowledge of early life.
There can be no mistake as to the prolific development of life in Cambrian seas, for fossils of this age are to be found in many parts of the world, where ancient sea bottoms now form part of the land surface. Invertebrate animals appear to have made much progress, but plants were either scarce or too small and delicate to be productive of fossils. It is probable, however, that seaweeds and other algae were flourishing along with the invertebrates, because animal life is directly or indirectly dependent on the existence of plants. The latter sustain themselves by taking carbon and nitrogen from air, water, and soil, but animals must obtain their requirements by eating plants or eating each other. They cannot obtain what they need from the inorganic world without this help from the vegetable kingdom.
One group of animals stands out prominently above all its contemporaries. Known as the trilobites they were by far the most distinguished and most characteristic of Cambrian invertebrates. Trilobites inhabited the warmer seas of this period and several later ones, but were extinct by the end of the Paleozoic era. Hundreds of species have been described, most of them under four inches in length. Well-known distant relatives now living are the shrimps, and other crustaceans. The name Trilobite has reference to the three lobes which are apparent in the form of the upper surface, the central lobe forming a broad ridge extending along the back. Beneath the outer lobes on each side there was, during life, a row of short, jointed legs used for swimming and walking, but these delicate appendages are seldom preserved in the fossils.
Second in importance among the animals of the period were the brachiopods or lamp-shells, not true mollusks although they were provided with similar shells composed of calcium phosphate or calcium carbonate. Shells are of two parts (bivalved) as in the case of clams, but the valves are above and beneath the body instead of on the right and left sides, which is the arrangement among mollusks. Although abundant as individuals, there were only a few species during the earlier part of the period; the number of species increased, however, and the race became very persistent. About seven thousand species have been described, and the race is not yet extinct although the number of living species is relatively small.
Cambrian life evidently included representatives of all the great divisions of invertebrates; sponges, jelly-fishes, worms, and primitive corals have been reported. At the end of the period there was an elaborate molluscan fauna. The closing of the period in North America was apparently a gentle elevation of continental areas and a consequent withdrawal of the sea.
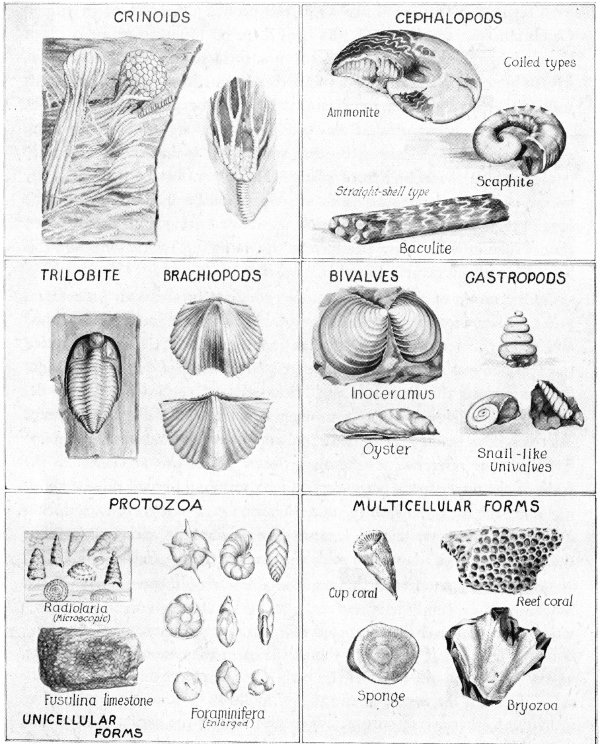
Invertebrate Fossils
Only a few prominent types have been selected from thousands of invertebrates known to zoologists. The forms illustrated are of frequent occurrence as fossils.
Extensive land areas must have subsided again early in the Ordovician period for marine sediments were laid down over a large part of the North American interior, and three epochs or subdivisions of the period have been based on as many invasions of the sea. In these ancient deposits the record of life continues to show new forms. Nothing of a very spectacular sort is recorded other than a great increase in the number of species among types that were established in earlier periods.
Trilobites were at their best, brachiopods continued to flourish, and the mollusks made new progress, especially the cephalopods, a group which includes our cuttle fishes and squids. Some of these predatory creatures attained large size and were no doubt masters of the sea. Typical forms were provided with tapering chambered shells that occasionally reached a length of twelve or more feet. Most of the shells were straight and trumpet-like or but slightly curved. Some were closely coiled and in this respect more like the well-known nautilus now in existence.
The bryozoans became very common in the later part of the period and corals made slight advances. Somewhat of a novelty at this time were the crinoids, commonly known as “stone lilies” although not plants at all. They have been described as starfishes with back turned downward and a thick stem attached beneath. Where they lived in great abundance the limestone deposits may consist almost entirely of their stems. Crinoids continued to produce a variety of forms throughout several of the succeeding periods.
The brachiopods were commonest of all animals representing this period, however, and their wide distribution, together with their known preference for warm waters, is taken to be an indication of mild temperatures prevailing over a large portion of the earth. Land plants are indicated by spore-bearing forms related to the ferns and mosses. Impressions of such plants have been found in Europe but, since most of the known rocks of this age were formed in seas, the marine algae are more abundant as fossils.
In the Colorado area, rocks of Ordovician time are exposed only in mountainous areas where they have been lifted high above their original levels. They are not especially rich in fossils although they have produced some fish remains which are of interest in that they suggest an age of vertebrates which is just ahead.
Since land floras and faunas had not yet become conspicuous the fossil record for this period is limited to those areas which were invaded by the sea. Apparently there was no such invasion of the present Colorado region, for rocks of this age are not in evidence. If they exist at all they are restricted to localized districts which are deeply buried under sediments of later periods. There may have been no Silurian deposition in this area, or such rocks may have been produced only to be destroyed by elevation and consequent subjection to weathering and erosion during a long interval of time, in which they were completely removed. In the region of the Colorado Rockies there is no evidence of returning seas until late Devonian time.
In other parts of the world, however, there was extensive deposition of rock-making sediments in seas which were inhabited by algae and invertebrates of the types previously described. Among the common animals of the time there were still numerous species of brachiopods, trilobites, corals, crinoids, and bryozoans. In addition to the primitive cone-shaped, cup corals there were several advanced types but the habit of building large reefs was not yet established.
“Sea scorpions,” really large crustaceans, flourished during Silurian time, and late in the period there appeared a race of true scorpions which lived on dry land or between high and low tides along the seashore. These were smaller and much like modern descendants but probably they did not wander far from the ocean shores where an abundance of food was available. These little scorpions, the largest measuring only two and a half inches in length, are the oldest air-breathing land animals of the fossil record.
It was not until the period was well advanced that fishes became numerous, and much of our knowledge of the beginning of an “Age of Fishes” has been obtained from European fossils. Although fishes are classed with the vertebrate or backboned animals there are large groups which do not have bony skeletons but are provided instead with a simple framework of cartilage. Among the earlier and more primitive types were the ostracoderms or bony-skinned fishes with no internal bones and only a small amount of bony substance in the armor-like plates and scales which covered the forward portion of the body.
The ostracoderms comprise a small group of fishes about which very little is known. They appear to have been inhabitants of fresh-water streams as well as lagoons bordering the seas, and may have been related to the small sharks of the time. They lived during the Ordovician, Silurian, and Devonian periods, and left no descendants now recognized 37 among living creatures. A much larger type of armored fishes is known as the arthrodires, a name which refers to a pointed neck and an arrangement of the armor plates to permit a movement of the head. These were the most ferocious fishes of the Silurian and Devonian seas, some of them reaching a length of twenty feet though most were much smaller. Their jaws were provided with formidable shearing and crushing plates instead of teeth.
The Devonian is one of the most outstanding of all periods from the viewpoint of life development. Dominance of the fishes is its greatest achievement, the invertebrates remaining about as they were and the higher vertebrates barely in evidence, but life on a large scale was no longer confined to the seas. Fresh-water fishes became prominent and land plants well established. The first forests appeared, with fern-like plants predominating although woody trees of several types and considerable size were included. It is quite possible that extensive land areas had been well supplied with vegetation during earlier times, but the delicate tissues of plants are far less likely to be preserved than the limy parts of animals. The fossil record, therefore, cannot be expected to reveal more than a suggestion of the progress made at this level of living. The story of plant life becomes much clearer in the next period when conditions were more favorable for the production and preservation of plant fossils.
Land animals of the time are almost unknown. A few snails and scorpions have been found, and some footprints made by early amphibians. Insects probably were in existence although the evidence is not quite clear on this point. The increasing number of fresh-water fishes, however, may be regarded as a sure indication that inland conditions were becoming more favorable for plant and animal inhabitants of all kinds.
The extent of development among the fishes cannot be accurately indicated by naming a few types, for it is mainly in the number of species and genera within the larger groups that progress is seen. In general it may be stated that the fishes of the period had not yet acquired the bony skeleton and typical form of familiar modern species. Skeletons were of cartilage, partly hardened in some instances by lime. Armor plates were customary with certain races but were not present among all fishes. Neither were these armored forms exceptionally large, as compared with living sharks. Although occasional giants appeared, the majority were small. Many were sluggish creatures with poorly-developed jaws, living as scavengers on sea and stream bottoms. Tail fins were usually unbalanced as in the sharks, or pointed and rounded rather than evenly forked.

Modernized Types of Fishes from Eocene Shales of Southwestern Wyoming
The great tribe of true bony fishes, such as the cod and perch, which includes more than ninety percent of the fishes living today, was not yet in existence. About one-third of the many kinds of fishes then living were related to the sharks, a group which is relatively insignificant in recent years. Nearly one-fourth of the total belonged to a tribe of enamel-scaled fishes, now represented only by a few sturgeon and gar-pike.
Lung fishes have never been a large group but it is noteworthy that they have had existence since Middle Devonian time. Living members of the race, inhabitants of Africa and South America, make a practice of burrowing into the mud of stream channels during dry seasons and are provided with lungs which enable them to breathe air in the manner of higher vertebrates. They survive the complete drying-up of the streams and live for months without water. Other forms, with less development of lungs, frequent stagnant pools and come to the surface occasionally for a breath of air. All are provided with gills also, which enables them to obtain their oxygen as other fishes do. They are believed to be a connecting link between the fishes and the early amphibians. More accurately, perhaps, they should be regarded as holding an intermediate position without being directly ancestral to any higher type of vertebrate animal.
Still dominant among the invertebrates were the brachiopods, on the whole averaging a little larger in size, and otherwise indicating congenial times for that type of organism. They reached the peak of their development during this period. Trilobites were declining although a few new and strangely ornamented varieties made a brief appearance. Crinoids apparently found living conditions less favorable during Devonian time, but in a later era they again became prominent. Corals were favored only at times and in certain localities. Along with the crinoids they appear to have suffered from the presence of an unusual amount of mud in the waters of their customary habitats. Both had a preference for clear water as indicated by the absence of fossils from limestones containing more than a very small percentage of muddy sediments. Crustaceans, similar to the sea-scorpions and better known as eurypterids, became prominent among fresh-water animals. Some were unusually large for creatures of this class, lengths of several feet being recorded from fragments. Gastropod mollusks came into prominence in localities where living conditions were favorable. Bivalves continued to thrive but the 40 cephalopods had a rather meager development considering the heights they were to achieve in subsequent periods.
In western North America the large expanse of territory known as the Great Plains was evidently well above sea level during this entire period, for no beds of this age are found in eastern Colorado. West of the Front Range, however, there was some deposition of marine sediments during late Devonian time. Formations of this age are exposed near Salida and Glenwood Springs, on the White River Plateau, and in the San Juan region.
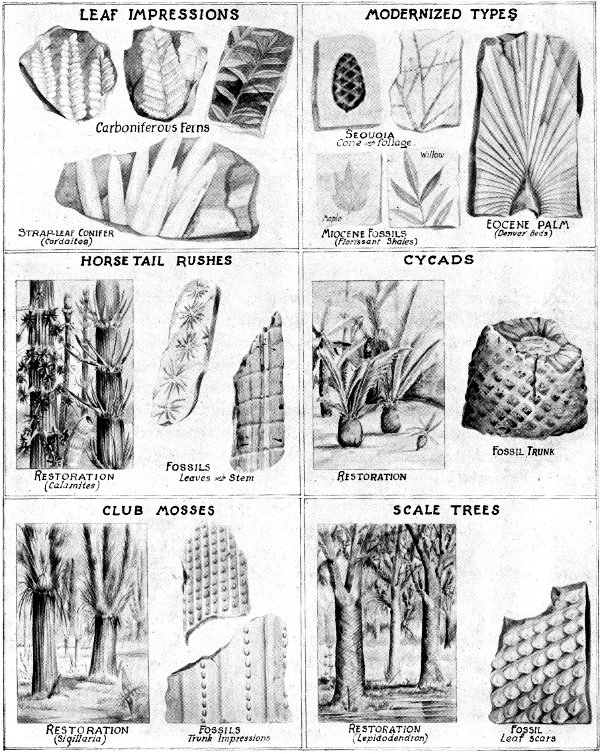
The Carboniferous period gets its name from the vast deposits of coal which were developed during that time in many parts of the northern hemisphere. Depressed land surfaces bordering the continents, and extending well into the interior of present boundaries, supported dense growths of vegetation and provided the swampy conditions most favorable to coal production. Varieties of plants which are now of small size and lowly position in the botanical world acquired the proportions of large trees.
Best-known fossils of the period are carbonized portions of the larger trees, and impressions left in the muds and sands of ancient bogs. Forest trees of several kinds reached the height of a hundred feet, with a trunk diameter of two to six feet. This size often is exceeded in modern forests, but by trees of an entirely different type. Considering the amount of development among the plants of earlier periods, Carboniferous forests provide an outstanding spectacle of advancing life.
Quite common among the larger trees were two varieties of club-mosses, also known as scale trees. They were cone-bearing evergreens with only slight resemblance to modern conifers. Instead of seeds they produced spores, a method of reproduction which is practiced among ferns. The trunks were marked from bottom to top with uniform patterns of cushions and scars indicating the points at which leaves were attached during the earlier stages of growth. In the Lepidodendrons the rows of scale-like cushions wind spirally upward while among the Sigillaria there is a vertical arrangement of leaf-scars which resemble the imprints of a seal, these impressions being in straight and parallel rows on a surface which may be either ribbed or smooth. The leaves of scale trees were stiff and slender, and arranged in grass-like tufts at the top.
Calamites, related to our horsetail rushes, were somewhat smaller than the scale trees. Their trunks consisted of a thin, woody cylinder with a pithy interior, and were marked at intervals by nodes which gave them the “jointed” appearance of a bamboo stem. Leaves were arranged in circles around the nodes of main stem or branches. Spore-bearing cones appeared at the tips of the stems.
Prehistoric Plants
Some of the larger and better known plants of past ages are shown as reconstructed by artists. Finer details of the reconstructions often have to be neglected because of uncertainties due to the scattered and fragmentary character of the fossil record.
Also included among the larger trees were the Cordaites or large-leaved evergreens, tall and slender, seed-bearing but not true conifers as yet. Leaves were strap-shaped or grass-like, the larger ones having a length of six feet and a width of six inches. Trunks were woody, resembling pine, but with a central pith. The flowers were small and resembled catkins in form.
Ferns and fernlike plants were so numerous that the period has been known as an age of ferns. Earlier knowledge of these forests was based on fossils of a fragmentary character from which an accurate association of parts could rarely be obtained. A general relationship with the ferns was apparent, but careful study of additional material has given us a rather different view of Carboniferous plant life and we note a highly diversified array of forms with many suggestions of modern tendencies. The flora as a whole may be regarded as highly specialized for the conditions which prevailed at the time and were not to continue through subsequent periods. Warm temperatures and abundant moisture were essential especially to spore-bearing types, and the cold, arid conditions of the next period put an end to many of the groups, or greatly reduced their prominence.
This could be regarded equally well as an age of insects, for some of these invertebrates acquired the greatest size they have ever had, particularly the dragonflies with a wing-spread of more than two feet in one of the largest fossils so far discovered. Cockroaches numbering upwards of five hundred species have been named. Though large they are hardly to be regarded as giants, lengths of three or four inches being about the limit.
Some of the insect types of today quite evidently existed among the inhabitants of Carboniferous forests, but it is apparent that there were also some antiquated forms which may have descended from the trilobites. Although some authorities regard this as the period in which insects originated, there are others who maintain that definite beginnings are not established so readily on present evidence. Spiders are believed to have made their appearance at this time.
Four-footed vertebrates resembling salamanders were prominent animals of the Carboniferous swamps. At first adapted to a life in water and later to land conditions, they are known as amphibians, the name being based on the ability to live in two different kinds of environment. Common living representatives of this group are the toads and 43 frogs, but these tailless forms are not known among fossils of the Paleozoic era and are almost unknown throughout the Mesozoic. The Age of Amphibians, as we apply that phrase, was definitely not an age of toads and frogs.
These primitive land animals were of different types, ranging from much smaller sizes up to the length of a crocodile. Most of them had short legs, and feet which were suitable for locomotion upon land, but many of the creatures probably spent most of their lives in the water. Tails were usually high and flattened as if for swimming, sometimes long, at other times greatly shortened in proportion to the body. Heads were generally large, jaws long, and mouths wide.
Before the close of the period true reptiles appear, but this race of animals is destined to make a more spectacular advance than the amphibians and will be discussed in connection with Mesozoic life. The amphibians, however, are regarded as being the ancestors of the reptiles as well as the higher quadrupeds which follow them. Although living reptiles are readily distinguished from living amphibians there is a different situation with regard to these primitive forms, for among the fossils it becomes increasingly difficult to separate the two groups as new material is investigated.
Invertebrates had their ups and downs during the period. Trilobites became scarce, and brachiopods for a time were the most abundant of the shelled animals but later declined rapidly. In favorable localities the crinoids established a wonderful record for new species before the period had advanced very far. Hundreds of species of Carboniferous invertebrates are known, and in many of the rocks of the period they are the only fossils to be found, for the vertebrates were still unable to venture far from the swampy districts, and much inland territory was too well drained to support either the floras or faunas then existing.
In the Colorado area there are both marine and continental formations but the great coal-making forests and their inhabitants were limited to other localities. As a consequence this region is not famous for Carboniferous fossils.
For a time there was no great change in North America following the opening of the Permian. Then began a series of mountain-making movements and continental uplifts which drained the swamps, lakes, and inland seas. With the passing of the vegetation which had established itself in and around these areas much of the animal life followed. It is 44 probable that a considerable proportion of the marine life survived, much more than is indicated by the fossil record, but the receding seas carried the survivors into territory which is now inaccessible to fossil hunters.
After Middle Permian time the climate everywhere seems to have been cold and dry. By the end of the period there had been accomplished more geographical change throughout the world than at any time since the beginning of the Paleozoic era. Traces of the crustal movements which produced new mountain ranges can be followed in Europe, Asia, and North America. The Appalachian region was raised to a great height, possibly in excess of three miles. A major disturbance of this character is known among geologists as a revolution, and to this particular one the name “Appalachian Revolution” has been given.
The elevation of continents necessarily changes their coast lines. This, in turn, influences ocean currents which have an important bearing on climatic conditions. In addition to this, the elevation of mountain systems not only rearranges the distribution of hot and cold winds over the land areas but it may produce barriers to the migrations of floras and faunas, confining them to areas in which it is no longer possible to live. When the effect upon plants and animals is considered, it is easier to understand why a line is drawn across the geological time chart at such a point and an era of prehistory is regarded as closed.
During the Permian period there was recorded in the rocks more widespread glacial action than ever before or since. With less inland water to provide the necessary evaporation there was a marked decrease in rainfall, and arid or semi-arid conditions replaced the hospitable climate that had been such an important factor in the prolific life of the Carboniferous. The struggle for existence became intense, but hardier types of plants and animals, with greater ability to adapt themselves to adverse conditions, established themselves here and there, as ancestral forms became extinct. Most of the large spore-bearing trees died out and seed-producing varieties began to acquire prominence, among them the coniferous evergreens. Ferns, however, proved their adaptability by producing some new forms which became prominent in Permian floras.
The prehistoric amphibians have been divided into three orders, one of which includes all the larger forms. This group, known as the labyrinthodonts, continued on through Permian time but began to show backward tendencies, with dwindling limbs and a return to life in the water. Among the larger land varieties are typical fossils ranging from about fifteen inches to five feet in length. In outward appearance they differed from Carboniferous amphibians. One of the other orders, including a great diversity of smaller forms, became extinct during this 45 period, leaving no known descendants. The third order is regarded as the oldest, and probably the ancestral group from which the modern newts and salamanders originated.
The most successful of Permian land animals were the peculiar reptiles that learned to live in drier regions. Like the horned toad and Gila monster of our arid southwestern United States, the larger Permian reptiles were four-footed animals. In size and shape they were not greatly different from amphibians then living. An exception to this rule, among some of them, is the development of long, bony spines above the vertebrae of the back. A fairly common fossil of this type, found in Texas and known as Dimetrodon, had a total length of six feet, about half of this being in the tail. The tips of the spines adorning the back reached a height of three feet or more and there was probably a covering of skin over these bones, which would produce a sail-like structure or “fin” of large size. Its use has not been explained but it provides an easy name for these odd creatures—the “fin-back lizards.”
Rock deposits produced in arid regions usually have characters which are not difficult to recognize. Gritty texture, irregular bedding, red color, and gypsum are common features. Formations of Permian age are to be found in Colorado but better fossil deposits have been discovered in Kansas and Texas.

Marine Reptiles: Plesiosaur (Lower Skeleton) and Mosasaur
The Mesozoic Era produced many types of reptiles besides the dinosaurs. Two of the marine forms are shown in this illustration, both from Cretaceous beds of the western United States. Plesiosaurs were the giants of the seas in their time, lengths of forty to fifty feet being not uncommon. A long flat tail provided locomotion for the mosasaurs whereas the plesiosaurs resorted to the peculiar limb structures known as flippers or paddles.
The Mesozoic, or era of middle life, was a long stretch of time during which there was marvelous development among the reptiles. Many strange types were produced and most of them became extinct before the end of the era. The reptilian stock branched out in many directions. Types emerged which differed from one another so widely that their mutual relationships have become obscure. Hideous and fantastic creatures suggesting sea serpents and dragons were worldwide in distribution. Reptiles of the air and seas acquired large size and weird forms, but greater advances were made upon land.
The flying reptiles or pterosaurs flourished in Jurassic times with some of the larger varieties surviving until near the close of the Cretaceous. Although these winged lizards were the first of the vertebrates to fly they are not to be confused with birds. They were without feathers, and the earlier forms were provided with long tails bearing a flattened rudder-like tip. One of the best known of this type had a length of about eighteen inches. Its jaws were long and provided with sharp teeth. The wings were membranes attached to body and legs, stretched and manipulated by means of greatly elongated fingers. In later types there was a reduction in tooth equipment and length of tail. Pteranodon, found in Kansas, had a wing spread of twenty-five feet, a large toothless beak, a short body, and a mere stub of a tail. It was one of the last of these winged monsters.
Several types of marine reptiles appeared during this era, among them the plesiosaurs which first appeared in Triassic seas. These peculiar animals were serpent-like with regard to the character of head, neck, and tail, but in other respects were quite different, the short barrel-shaped body being provided with four large paddles corresponding to the usual limbs of quadrupeds. Fossil remains of these animals are common in many Jurassic and Cretaceous deposits, some of the largest exceeding forty feet in length. Mosasaurs, also marine carnivores, inhabited shallow Cretaceous seas throughout the world and are especially abundant as fossils in the Kansas chalk beds. These were elongated forms with a resemblance to salamanders in some respects but provided with long pointed jaws and sharp teeth. Swimming was accomplished largely by the tail though probably aided to some extent by four webbed paddles or flippers. The ichthyosaurs were more fish-like in construction, as the name implies. The limbs were short and broad, and there was usually present a well-developed tail-fin as well as a large fin on the back. They were especially abundant in Jurassic time. Fossils are fairly common in marine deposits 48 of western North America. Mosasaurs and ichthyosaurs were about half as long as the plesiosaurs.
Most spectacular of the prehistoric reptiles were the dinosaurs, a large group of animals varying greatly as to size, form, and habits. They were adapted for a life on land though many of them probably spent much of their time partly submerged in the waters of lakes and streams. There is little that can be said of the group as a whole other than that all of them were reptiles. Further than that it is necessary to regard them as belonging to several different subdivisions of the Reptilia. Classification has been difficult and the names used for the various subdivisions are often misleading to the layman who tries to understand the terminology.
Ancestral reptiles were five-toed and five-fingered but among the dinosaurs there were many departures from the standard formula. Three or four of the digits were commonly well developed, the others when present being shortened or reduced to mere rudiments. Early in the history of dinosaurs there was a division of the stock into two main branches, each of which includes a variety of types and sizes, and is again subdivided. The two main groups are best recognized by the construction of the bony framework which comprises the pelvic girdle or hip region of the skeleton. In order to avoid technical difficulties, however, the remaining discussion of these interesting reptiles will be confined to a few names and descriptions which serve to illustrate roughly the great amount of variation that developed from the comparatively simple ancestral pattern. The plan according to which the dinosaurs are usually classified is barely suggested by the types described.
The meat-eaters were active creatures provided with powerful jaws and teeth. They were unarmored, moved about on their hind feet, and during their time were the most highly advanced of all animals. Tyrannosaurus with a length of forty-five feet or more, and Deinodon, nearly as large, were among the greatest of these. Both lived in the Cretaceous period. Their teeth were simple but strong, knife-like, curved, and finely serrated. Skulls were large and the forelimbs were reduced almost to a state of uselessness. Large carnivores lived also during Jurassic time and even as far back as late Triassic. Early Triassic forms were of smaller size.
More primitive flesh-eating dinosaurs of the Triassic and Jurassic periods were delicately proportioned and lightly built bipeds bearing some resemblance to birds. Struthiomimus, which means ostrich-resembling, was about the size of the bird which provides the name. It was slender in the limbs, three-toed, long necked, long tailed. The skull was small, forelegs long for a biped. Unlike most dinosaurs it was toothless. All these bird-like carnivores were small as compared with other contemporary forms. Compsognathus, of Germany, and one of the smallest of all dinosaurs, had a length of less than three feet, including the long tail.
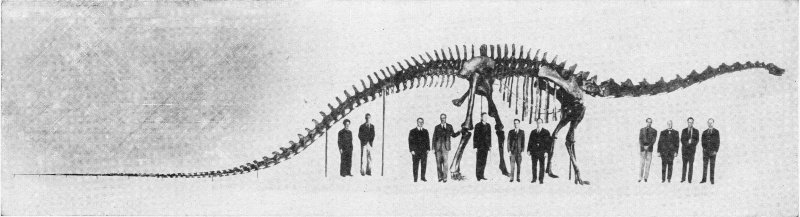
One of the Large Jurassic Dinosaurs (Diplodocus longus)
This magnificent specimen, exhibited by the Denver Museum of Natural History, has a length of seventy-five feet six inches. Two years were required to complete the task of removing the bones from the matrix rock and preparing them for mounting. Diplodocus was one of many large reptiles which inhabited western North America a hundred and fifty million years ago. The skeleton was obtained from the Morrison beds of eastern Utah. The same formation is exposed in many Colorado localities, including the foothills west of Denver, where it acquired its name from the town of Morrison.
In Jurassic time there became prominent a group of large dinosaurs which were more equally developed as to fore and hind limbs. They were sluggish creatures, quadrupedal in their manner of locomotion, vegetarians in regard to their diet. Some of them reached enormous proportions and it is believed that they resorted to life in the water in order to get part of the weight off their feet. Diplodocus and Brontosaurus are the names of well-known giants in this group. They had long necks and tails, very small skulls, were the largest of all land animals and are known to have reached a length of eighty feet or more. Some estimates, based on measurements of incomplete skeletons, have exceeded one hundred feet, but these extremes are somewhat questionable. Diplodocus was the more elongated of the two, with much of its length in the whip-like tail. Our mounted skeleton has a length of seventy-five feet six inches, measured along the vertebrae. Its height at the pelvis is twelve feet six inches.
The teeth of these large quadrupeds are of a slightly broadened and blunted form which has caused some speculation as to their possible use. It has even been suggested that the animals were fish-eaters but this seems impossible in view of the great size and general characteristics of the group. Although they differ extremely in some respects, they are regarded as being more closely related to the carnivores than to the herbivores of the second great branch of the tribe.
The unquestioned herbivores, constituting this second branch of the dinosaurian race, also include both bipeds and quadrupeds. The better known plant-eaters were large animals but not such monsters as Tyrannosaurus or Brontosaurus. Of the bipeds, Trachodon is perhaps best known. It is one of the duck-billed dinosaurs which had an average length of about thirty feet. The duckbills were unarmored, active animals, good swimmers as well as runners. They were prominent and widely distributed during late Cretaceous time. Many skeletons have been found in western North America. Natural casts and impressions of mummified remains indicate that the hides were scaly and the feet provided with webs between the toes. The bill was broad, flat, and toothless, but the sides of the mouth were provided with a large number of simple teeth closely arranged in parallel rows. The fine skeleton exhibited in our hall is thirty feet six inches in length. Near relatives of Trachodon, such as Corythosaurus had hollow, bony crests, combs, or tubular structures on top of the head. These may have been of some service in connection with breathing while feeding under water.

A Duck-billed Dinosaur of the Cretaceous Period (Trachodon mirabilis)

Stegosaur (Stegosaurus stenops)
Among the quadrupedal vegetarians an interesting family is represented by Stegosaurus, a late Jurassic dinosaur having a length of about twenty feet. These creatures had heavy limbs, all used in walking, an arched back, and almost no brain at all. A double row of large flattened plates standing upright and extending from the rear of the skull nearly to the tip of the tail provided some protection for the back of the animal, but otherwise there was no defensive armor. Several long spikes at the end of the tail probably served as weapons. The mounted skeleton in our collection was obtained from Garden Park, near Canon City, Colorado, a district which has long been famous for dinosaur remains.
The ankylosaurs were more completely armored with closely set bony plates fitting neatly over the body. They were of about the same size as the stegosaurs but the body was broad and somewhat flattened. These armored quadrupeds apparently lived only during the Cretaceous period, after the disappearance of the stegosaurs. Their tooth equipment was very poor and in a few cases entirely lacking. Ankylosaurus and Nodosaurus are good examples of the type. They have been described as animated tanks and are sometimes referred to as having the appearance of enormous horned toads.
Among the last of the dinosaurs to come and go were the horned quadrupeds known as the Ceratopsia. Their entire history appears to have been confined to the Upper Cretaceous and the closing stages of the reptilian era in America. Triceratops and Monoclonius are well-known representatives of the group. Besides the horns, which appeared above the eyes or near the center of the nose, there was a broad, flattened, backward extension of some of the skull bones which produced a great frill or collar reaching over the neck as far back as the shoulders. This frill, combined with the large skull, gave the animal the appearance of being nearly one-third head. Triceratops had three horns, Monoclonius only one. The average length of the animals was slightly under twenty feet.
Although very little is known about the ancestry of the horned dinosaurs a valuable discovery in Mongolia may throw some light on the subject. A small dinosaur with a well-developed frill, but no horns, once inhabited the region of the present Gobi desert, and in recognition of the apparent relationship it has been named Protoceratops. In addition to numerous skeletons, several nests of eggs were found in association with the bones. Until this discovery was made, dinosaur eggs had been practically unknown. A reproduction of one of these nests is among our exhibits.

A Sea Turtle of Cretaceous Time (Protostega gigas)
This marine animal belongs to a group which became extinct near the close of the great reptilian era, but a few related forms still survive. Their weight is greatly reduced by the peculiar construction of the shell, and the front feet are enlarged for use as oars, an excellent illustration of the manner in which a land type can become adapted to life in the sea.
With the possible exception of a very few short-lived survivals dinosaurs were extinct before the opening of the Age of Mammals, many of them for millions of years. Along with them went other types of ancient reptiles, and the cause of their extinction is a problem which may never be solved. Conditions remained favorable for the turtles, which made their first appearance during Triassic time, and for the crocodiles, which date back to the Jurassic period. Snakes were only at the beginning of their history as the era closed. The survival of these modern forms suggests that they were favored to a greater extent than the dinosaurs during a prolonged period of changing conditions the full details of which are unknown to us.
In general it is to be expected that disaster would first overcome the highly specialized creatures, such as the dinosaurs, which had become more delicately adjusted to the particular environments in which they lived. It appears that some of them had been too progressive up to a certain point, but not sufficiently adaptable to get beyond that stage, or fortunate enough to make their advances in directions that could be followed, through fluctuations in the matter of food supply, predatory enemies, climate, and other factors which bear upon success and failure.
The reptilian era closed with exceptional volcanic activities in many parts of the world, but these cannot account for the disappearance of the highly diversified and abundant reptilian life. The eruptions were merely incidental to movements and readjustments in large masses of rock comprising the earth’s crust or surface. Such crustal folding and elevations always have been of serious consequences to both plants and animals because of their effect upon drainage and climate. There were disturbances of this kind in western North America in late Jurassic time, with folding and uplift in the region of the Sierras and probably extending from Mexico to southern Alaska. A great trough to the east of this elevated district was produced in the course of these movements and provided access to the sea from south to north. During the Cretaceous period there were repeated invasions and retreats of the sea by way of this great depression, consequent upon slight changes in the elevation of the floor. Hence there are numerous marine formations in Colorado and adjoining states, some of them rich in fossils.
Before the close of the Cretaceous period the sea had made its final departure from this region, and the Mesozoic era was terminated by revolutionary disturbances which brought about the uplifting of a new mountain system. The Rocky Mountains may be regarded as part of this system and to have had their birth at this time. The Rockies, however, show unmistakable signs of repeated elevation, with intervals of erosion 56 during which there was great reduction of their total height. What we see of them today is the result of more than fifty million years of continuous geological activity.
Some idea of the Mesozoic climate is obtained from the character and distribution of the plant life. Triassic floras are not large and there is very little fossil evidence for the earlier half of the period. It is quite possible that arid or desert conditions prevailed for a time in much of North America, as at the close of the Paleozoic era. Plant life was at first not abundant, and conditions were unfavorable for the production of fossils. In Upper Triassic rocks of Virginia, however, there are signs of swampy conditions, with rushes and ferns predominating. Adjoining forest areas were well timbered with large coniferous evergreens which show no annual growth rings, as similar trees do in regions where cold winters alternate with warm summers. This suggests, for that time and place at least, a uniformly warm climate, lacking seasonal variations. Warm temperature or subtropical climates are indicated again by some of the Jurassic and Cretaceous plants, but intervals of lower temperatures and variable climates are also apparent. Palms, figs, and other trees, very similar to modern types now living only in warmer regions, were widely distributed in late Cretaceous time, and their range was extended into regions which have since become too cold to support such growths.
The trend toward modern forms in the plant world was gradual, but throughout the era there were occasional novelties that attract the attention of botanists. Ferns and horsetail rushes, reminiscent of the Paleozoic forests, soon began to lose their prominence as the seed-bearing trees gained the ascendency. Mesozoic time could well be called the age of cycads, because of the striking performance of this plant group. Different varieties flourished in the three periods, with the Jurassic standing out as the time of greatest abundance.
To the uninitiated, the usual cycad fossils resemble “petrified pineapples,” but these are merely the scarred stems or trunks of small to medium-sized trees with a tufted arrangement of leaves at the top, and usually without branches. Foliage and habit of growth suggest something more like large ferns or low-growing palms, with short, thick trunks seldom more than fifteen feet tall and many of them under three feet. The leaves are rarely found entire or attached to the trunks, but occasional discoveries indicate a leaf-length of about ten feet. Although they are classed among the first and lowest of seed-bearing plants, and 57 in this respect are related to the conifers, their appearance was quite unlike that of the modern cone-bearing evergreens.
More nearly resembling the common conifers of today were the sequoias, of early Mesozoic origin and far more abundant during Cretaceous time than they are at present. The maidenhair trees, now represented by a single species of Ginkgo which is cultivated principally in China and Japan, were never very prominent but are of interest as an ancient family that persisted throughout the Mesozoic and down to our own time. Before the close of the Cretaceous period the flowering plants had greatly outnumbered the spore-bearing groups, such as the ferns and horsetails which were formerly so abundant. We know little of early flowers, however, except in connection with trees, the large gayly colored blossoms of the type now conspicuous in woodlands, meadows, and gardens being later arrivals and poor subjects for preservation as fossils.
Cretaceous floras were surprisingly modern in character, far in advance of the animal life. Poplars, plane trees, magnolias, palms, figs, oaks, and buckthorns were abundant at the close of the Cretaceous, as indicated by fossils of the Laramie formation, which is the surface rock in many localities near Denver. Also abundant in various places at this time were walnut, hazelnut, laurel, tulip, maple, beech, birch, breadfruit, ivy, holly, and many other well-known trees and shrubs. Sedges and grasses, which became so important to the herbivorous mammals of the next era, made their first appearance in Cretaceous time but were then inconspicuous.
The abundance of plant life in the Colorado area during the Cretaceous period is indicated by the extent of coal deposits of this age. About one-fourth the area of the state is underlain by coal seams varying in thickness from a few inches to fifty feet or more, most of it being Cretaceous. In the northern Colorado district the coal-bearing formation is the Laramie. Near Denver there is some coal in the Arapahoe formation which overlies the Laramie and is of later age.
Coal mines often produce excellent plant fossils, and occasionally other evidence of prehistoric life. In a mine near Canon City, Colorado, a series of natural casts of dinosaur feet was taken from the overlying rock after the coal had been removed. One of these, in the Denver Museum of Natural History, is seen to consist of sandstone inside a very thin layer of dark clay. Flattened against the lower surface is the carbonized stem of a Cretaceous plant which grew in the swamp where the coal deposit was formed.
Since the shape of dinosaur feet is unmistakable we can only assume 58 that a large reptile of this type walked over the surface of swampy ground in which a great thickness of decaying vegetation had accumulated. A layer of mud settled over the top and became sufficiently firm to retain the mold of the feet as the animal moved along. Any plant material either on the mud or included in it was pushed to the bottom of the impressions and flattened out by the weight of the huge creature. Then sand was washed into the footprints from some nearby source during a heavy rainstorm.
Following these events there was probably a subsidence of the area, and a great thickness of rock-making sediments was built over the ancient swamp. The buried vegetation gradually became converted into coal, the sand consolidated into a firm sandstone, and the mud produced the shales forming the roof of the present mine, which is now at an elevation of a mile above sea level as a consequence of the general uplifting of the Rocky Mountain region during late Cretaceous and subsequent time.
When the coal was removed, the hard sandstone casts separated readily from the softer shales surrounding them. A small amount of the shale adheres to the sandstone, and some of the flattened vegetation, now in the condition of coal, still remains attached.
As in other eras, the invertebrates fluctuate with the periods. Characteristic forms appear, become more or less prominent, then in many cases decline or disappear. Variations among the mollusks are particularly helpful in the identification of rocks which originated in the Cretaceous seas. Clam-like bivalves of the genus Inoceramus, the straight-shelled ammonids known as Baculites, and oysters, are locally common in some of the formations exposed a few miles west of Denver.
The ammonids, or “ammonites,” were extremely abundant throughout the world during Mesozoic time. Their shells were chambered like those of the pearly nautilus, a related cephalopod mollusk inhabiting tropical seas at the present time. While only four species of the Nautilus tribe are living today, thousands of species of ammonids swarmed the prehistoric seas. Many new forms came into existence in Triassic time but few survived the period. A pronounced revival occurred in the Jurassic, only to be followed by a decline and eventual extinction at the close of the Cretaceous. Ammonites measuring three or four inches across the diameter of the coiled shell were about average size, but diameters up to three or four feet were not uncommon. Externally the shells were ornamented with ribs, knobs, and spines; inside was a pearly 59 lining. The partitions were thin and composed of the same pearl-like substance as the lining. Each partition becomes wavy as it approaches the shell, and the line of union has a distinct pattern which is seen in specimens which have lost the outer shell layer. This wavy suture line becomes more complicated in the later members of the race, and the peculiar markings developed by the repeated partitions provide a convenient method of identification.
The belemnites or ink-fishes, regarded as ancestors of the cuttlefishes now living, comprise another group of carnivorous mollusks. These, however, had lost the external shell, and the usual fossil is part of an internal shell or “skeleton,” known as the guard. This limy structure has the form of a pointed cigar, and is seldom over a foot long although the total body-length of the larger animals was commonly about six to eight feet. Several hundred species have been described, the majority being of Jurassic age. They declined rapidly toward the close of the era.
The invertebrate life of the Mesozoic was strongly dominated by mollusks, with cephalopods in first place, the bivalve pelecypods and the single-valve gastropods or snail-like forms sharing subordinate positions. The dominating trilobites, sea-scorpions, and tetracorals of the Paleozoic had disappeared, while the brachiopods and crinoids were greatly modified and more like the forms which live today.
Crinoids became moderately abundant at various times, but in many ways different from their relatives of the preceding era. Some of the largest known species, with stems estimated as fifty feet long, have been found in lower Jurassic rocks. A great abundance of microscopic life is indicated by the frequent occurrence of chalk in the Cretaceous formations. Corals of the modern reef-building type (hexacorals) were common in the warm seas of a large part of the world.
The insects of the early Mesozoic are represented by few fossils although it is evident some new forms were becoming established. The warm climates prevailing throughout much of the world appear to have been a favorable factor in the progress of insect life. In addition to the older cockroach and dragon-fly types may be mentioned the arrival of grasshoppers, cicadas, caddis-flies, beetles, and ants.
Several hundred species are found in Jurassic rocks, and by the end of the Cretaceous period most of the insect families now known to us were probably in existence. The record is seriously obscured by the erosion of rocks which so frequently marks the end of a period, also by the small size of the subjects, and by the exceptional conditions required for the production of such delicate fossils. Among the last of the familiar insects to appear were the bees and butterflies. These evidently 60 came in with the more advanced types of flowering plants that produce the nectar on which many insects feed. It is probable, too, that without the arrival of these insects and their service in the pollination of flowers, the floras of today would be rather different from what they are.
It is not surprising to find that birds made their first appearance in the Mesozoic era, for of all animals they are most like the reptiles as a class. Feathers are about the only dependable characteristic of the entire group, nearly every other feature being matched by some reptilian creature of great antiquity. The nesting habit, which includes care of the young as well as the eggs, is a matter of progress which relates to flight and to warmer body temperature. It appears to have been developed by forest-dwelling types living among trees and nesting there in comparative safety from enemies prowling on the ground.
The oldest known prehistoric birds were found in lithographic stone of Upper Jurassic age. Archaeopterix was discovered in 1861 at Solenhofen, Germany. Sixteen years later a similar bird in a better state of preservation was found in Bavaria. The latter was named Archaeornis. These Jurassic fossils are regarded as true birds by some authorities, while others believe them to be more nearly related to the reptiles, the opinions being based on careful studies of the skulls and other skeletal features. Both birds had teeth of reptilian character, and it is evident that there was no beak, for the jaws were covered with scaly skin. The bony construction of the long tail would suggest lizards rather than birds, were it not for the presence of feathers which were attached at each side. Head, neck, and parts of the body were covered with scales. Wings were well provided with stout feathers but the skeletal framework indicates that the birds were gliders rather than true flyers. Claws on the wings served like fingers to aid in climbing among the branches of trees, a practice which is occasionally noted among the young of living birds. In adult birds of today, however, the claw-like appendages of the fore limbs are greatly reduced and of little service.
The next fossil birds of importance have been found in Cretaceous rocks of Kansas, both of them fishers of the seas instead of forest inhabitants. Ichthyornis was a small bird, standing about eight inches in height, a powerful flyer with reptilian jaws and teeth. Hesperornis was built for diving and swimming, like the loon, but was somewhat larger and provided with teeth. Its wings were too poorly developed to be of use in flying.
Toothed birds became extinct with the close of Cretaceous time, 61 and the ancestors of modern types were in existence before the Age of Mammals, but fossil remains are few and poorly preserved. Large ostrich-like birds, however, are known to have lived in North America during the Eocene period. One of these, named Diatryma, stands nearly seven feet tall in the reconstructed skeleton. Its legs are heavy, wings greatly reduced, beak massive. In its relation to modern birds it is possibly nearer to the cranes than the ostriches.
Flightless birds of large size are known from many parts of the world and seem to have been prominent throughout the Cenozoic era, as they are today in the southern hemisphere. Aepyornis lived in Madagascar during the Pleistocene period and may have become extinct quite recently. Its eggs are the largest known among fossils, several times the size of an ostrich egg. Also in this period the moas were living in New Zealand where their remains are still abundant. One of the largest, known as Dinornis, had about the same form as Diatryma but the neck was longer, head and beak smaller, legs better fitted for running, height about eleven feet.
A much smaller flightless bird, the dodo, became extinct in modern time. This former inhabitant of Mauritius and other islands of the Indian Ocean was related to the doves and pigeons, and had lost its power of flight through disuse of the wings. It was a clumsy, defenseless bird weighing possibly as much as fifty pounds. Actual remains are few and incomplete, and descriptions published by the explorers who knew the bird two centuries ago are not entirely trustworthy. In the Pleistocene Rancho la Brea beds of California the largest of all prehistoric flying birds has been found, a vulture bearing the name of Teratornis. Re-assembled skeletons show them to be slightly bigger than existing condors.
The monotremes or egg-laying types of mammals such as the duck-bill and spiny anteaters which now inhabit Australia are almost unknown as fossils. Marsupials, the next higher living group, which includes the opossum and kangaroo, appeared at the end of Cretaceous time along with the placentals or higher mammals which dominate the history of the Cenozoic era. Nevertheless, there are a few teeth and jaws from rocks of Triassic and Jurassic age to indicate that small mammals, from the size of mice to slightly larger than rats, existed throughout most of the Age of Reptiles. There is no complete skeleton of any of the earlier forms, and little is known of their relationships either with living orders of mammals or with probable ancestors among 62 the reptiles. The record becomes somewhat clearer toward the end of the era but it is obscured again by the great disturbances which followed.
Looking back among earlier land animals for the origin of the first mammalian stock it is necessary to go as far as Permian or even Carboniferous time. Reptiles then living had many structural features in common with mammals, and mammal-like forms continued to flourish until late in the Triassic. An interesting group of such animals, named therapsids, was one of the earliest reptilian stocks to appear, and is well known from fossils found in the Red Beds of Texas and New Mexico, in Europe, South Africa, and Asia. Quite a variety of types is included in this group, with many advances in dentition, and modifications of the skull, limbs and pelvic construction which strongly suggests a relationship to the mammals.

Murals Over Fossil Exhibits, Hall of Mammals
Top: Eocene; Protylopus, Tanyorhinus, Patriofelis, Uintatherium, Turtle, Crocodile, Eohippus.
Middle: Upper Oligocene; Mesohippus, Merycoidodon, Hoplophoneus, Metamynodon, Poebrotherium, Trigonias.
Bottom: Pliocene; Teleoceras, Turtle, Synthetoceras, Amebelodon, Teleoceras.
The striking feature of life development in the Cenozoic era is the great progress and expansion over the earth of the mammalian races. The division of the era into periods, however, was based largely on a study of fossil mollusks. In the Paris basin of France, it was noticed by the geologists of a century ago that the youngest of the sedimentary beds contained the greatest number of recent or still living species. Successively downward into the older beds the percentage of recent species decreased until there were practically no living species represented in the oldest rocks of the series. From the percentage of recent forms among prehistoric ones it was proposed that the following division be made: Eocene, meaning dawn of the recent; Miocene, meaning less recent; and Pliocene, meaning more recent. Sometime later it was suggested that another period be added, and to this was given the name Pleistocene, meaning most recent. In 1854, the older Miocene formations were segregated and referred to a newly provided Oligocene period, this name meaning little of the recent.
Early geologists grouped the rocks in three great divisions, applying the names Primary, Secondary, and Tertiary. To these was added afterwards the name Quaternary, which applied to the youngest formations of the earth. Only two of these terms remain in common use at present: it is a frequent practice to refer to the combined Eocene, Oligocene, Miocene and Pliocene periods as the “Tertiary” division of Cenozoic time; to the Pleistocene and Recent periods as the “Quaternary” division. The geology of some remote future may be clearer with regard to the full significance of this subdivision of the Age of Mammals into two parts. It may be that a great era was concluded at the end of Pliocene time as others have been concluded, by the usual earth disturbances and climatic changes and by the decline of animals once prominent in the faunas of the world. Events of such character have registered their occurrence but may eventually prove to have been a series of minor events not comparable with the revolutionary changes that terminated other great time divisions. The favored practice of including ourselves and our times in the Cenozoic is based on a trend of opinion which holds that no great era has been ended since the Age of Reptiles was concluded.

Skulls of the clumsy, six-horned uintathere and the early, hornless titanothere form part of this Eocene display. In the mural these animals and the little “three-toed” Eohippus, smallest of horses, are pictured with a contemporary turtle and crocodile.
The oldest of Eocene rocks show a great variety of mammals and a strange assortment of forms far in advance of the Mesozoic record. Ancestries and successive stages of development have been only partially worked out, though details have been better preserved for some of the groups than for others. Some oddly shaped creatures such as the uintatheres reached their full development in a relatively short time and passed out of the picture before the end of the period. These animals, represented in our collection by Uintacolotherium, acquired large bodies and many horns, but a peculiar tooth equipment fitted them for a special diet which apparently failed to be supplied in sufficient abundance at a critical time.
On the other hand, we find in this period the ancestors of more successful groups, some of which continue on into modern times. Only a few of these histories can be traced in a brief sketch, but in a general way it may be stated that the successful races had modest beginnings and that they developed very slowly into what they are now, by a process of adjusting themselves, or by becoming better adapted to new or previously unused conditions in their respective environments. In their early stages the various types had much in common; they were generalized, rather than specialized for any particular kind of existence. What they were fitted for is best indicated by their teeth and feet, though other structural features frequently contribute valuable information. The rise of mentality is indicated by skull capacities and the increasing development of the upper lobes of the brain, as revealed by casts taken from the interior of skulls.
The creodonts were the earliest and most primitive of the flesh-eating mammals or carnivores. Many of them were small of body and brain, and equipped with teeth that indicate a mixed or largely insectivorous diet, or possibly the habit of feeding on carrion. Although there was considerable variation among them, and some tendency toward specialization, there was little to suggest the coming of more progressive groups such as the cats and dogs, with teeth perfected for the tearing and cutting of flesh, and feet especially fitted for the life of hunters. The ancestry of the cats cannot be traced farther back than the Oligocene but it probably connects somewhere prior to that time with the creodonts.
Cynodictis, an Oligocene carnivore slightly under two feet in length is commonly regarded as a primitive dog, but its characters are so generalized that it probably differs but little from the ancestors of many other carnivores. The skeleton of this animal suggests a slender and flexible body like that of the weasel, with somewhat shortened limbs and a long tail. It lived in forested regions and was probably more or less of a tree dweller. The more advanced carnivores required longer legs, better adapted for running and overtaking the prey, which is the dog’s way of hunting, or for stalking and springing upon the quarry, which is the method of the cat.

Moropus (Moropus cooki)
Though its teeth clearly indicate a diet of plant material, this strange animal had claws on its toes, like the carnivores. It is probable that the claws were used in digging for roots, as indicated by the artist.
A prominent group of mammals today is that known as the ungulates, or hoofed animals, which includes the horses, cattle, deer, swine, rhinoceroses, tapirs, and other types both living and extinct. Their probable ancestors were the condylarths, primitive ungulates of the Eocene period. One of these, known as Phenacodus, serves well to illustrate the general character of the early hoofed mammals. It was about five and one-half feet long, rather large for its time, with long tail and short limbs, low elongated skull and small brain, very similar in many respects to the creodonts or ancestral carnivores. The teeth, however, were partially of the grinding type so essential to the welfare of plant feeders.
The condylarths were five-toed animals and evidently provided with small hoofs, but the more progressive ungulates soon lost one or more of the toes, and a division of the group into odd-toed and even-toed branches became firmly established. Consequently, the families of ungulates having one, three, or five toes are classed together as being closely related to one another, and those having two or four toes are segregated in a second lot. The odd-toed clan, known as perissodactyls, included such animals as the horses, rhinoceroses, tapirs, and titanotheres, each of these types being placed in a separate family. The even-toed clan has been treated in a similar way and named the artiodactyls. In this division are such families as swine, cattle, deer, camels, oreodonts, and others. The odd-toed group dominated among the larger animals of North America for a time but has been completely replaced by the even-toed division which is still flourishing, although some of the older families have become extinct.
Among the exhibits of the Denver Museum of Natural History may be seen complete skeletons of extinct horses, rhinoceroses, titanotheres, and chalicotheres representing the perissodactyls. Moropus was one of the chalicotheres, an exceptional family which never became very prominent although it had a prolonged history and persisted in Europe and Asia after its extinction in North America. The family is grouped with the ungulates because of many similarities found in the molar teeth, skulls, and other parts of the skeletons, but the toes were provided with claws instead of hoofs. The use of these claws is somewhat of a puzzle: possibly for defense against carnivorous enemies, for dragging down branches in order to obtain food, or for digging roots which may have been an important part of the diet.
Titanotheres are represented in our collection by the skeletons of the large, horned type which was the last of the race and destined to extinction by the middle of the Oligocene period. Smaller hornless varieties of Eocene time are illustrated by skulls. This family of ungulates had an unprogressive dental equipment, and a small brain in a flattened skull. The molar teeth readily distinguish the group from other ungulates and enable us to trace the relationship between earlier and later varieties. These teeth were of a type which is soon destroyed by wear, and it is evident that the animals survived only so long as their environment provided them with an abundance of soft vegetation.

Titanotheres of Oligocene Time
The name of these animals refers to the large size though they were greatly exceeded in bulk by the mastodons and mammoths of later periods. Ancestral titanotheres, dating back to the Eocene, were hornless animals of much smaller size. These splendid specimens were obtained in Weld County, Colorado.
The large assortment of rhinoceros material provides an idea of the great abundance and variety of forms in this family which was once prominent in North America but no longer among the inhabitants of that continent. Some of the mounted skeletons have been restored on one side to show how these animals appeared in the flesh.
Of the even-toed ungulates there are also several types illustrated by complete skeletons. Merycochoerus, the subject of one of our mounted groups, represents the oreodonts, a large family of mammals whose history begins with the Upper Eocene and ends in the Lower Pliocene. The oreodonts were small animals, rather pig-like in form and quite common in the western plains region shortly after the time of the titanotheres. Ancient swine are represented in our exhibits by two mounted skeletons which were obtained from northeastern Colorado, where the bones were found associated with rhinoceros and titanothere remains. Some of these animals were of very large proportions, and the entire family is commonly known as the “giant pigs.”
Camels and closely related forms were quite abundant in North America from early Oligocene to comparatively recent time. Numerous types were developed during the course of their history, some small and delicately formed, others tall and clumsy and much like the giraffe in structure. Parts of many of these creatures have been found but the only completely prepared skeletons in our collection are of the little gazelle-camel, Stenomylus, from Lower Miocene deposits in northwestern Nebraska. Pleistocene bisons are represented by several complete skeletons and numerous skulls and horncores, some of the species showing an extreme development in the length of horns. With two of the bison skeletons are shown prehistoric weapon points, found with the bones and indicating that these animals were hunted by primitive men at some time near the close of the Ice Age. The artifacts first discovered near Folsom, New Mexico, by field workers of our Museum, have become known to archeologists as Folsom points.
The past history of horses is well known from an abundance of fossil material, ranging in age from the Eocene down to the present. Modern horses have only one toe in each foot, but there are remnants of two additional toes which may be seen only in the bony structure underlying the skin. Most of their ancestral relatives were three-toed as far back as the Oligocene period. During Eocene time, however, there was a stage which may be regarded as four-toed although it was evidently a temporary condition, linking known horses with more remote forms having five toes.
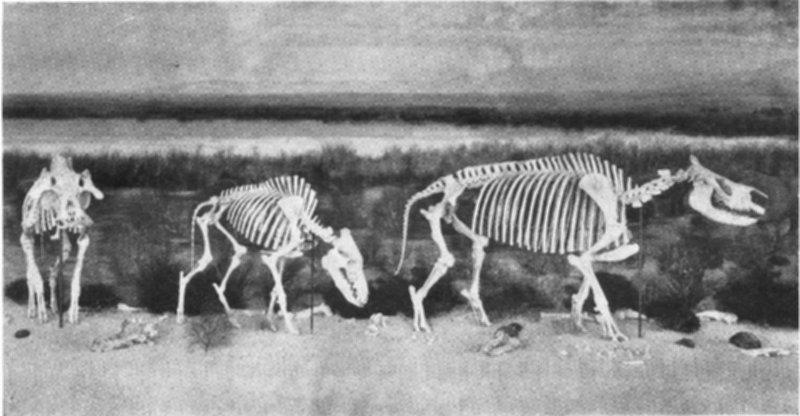
Oligocene Mammals From Weld County, Colorado
The giant pigs (Archaeotherium mortoni) at the left of the group, and the rhinoceros (Trigonias osborni) were common animals of the western plains region at one time.
Eohippus, the “dawn horse” as it has been called, is one of the oldest and best known of the American horses. Its relation to existing members of the family can be traced by means of changes in tooth structure as well as in the gradual reduction in the number of toes that is seen among intermediate forms. Its ancestors some day may be positively identified in that group of generalized, primitive, five-toed, hoofed mammals which are known to have lived at the beginning of the mammalian era, but such identification has not yet been established. Even Eohippus bore little resemblance to the familiar horse of today. Its height was only eleven inches, and in body form it had much of the appearance of a modern dog. There were four toes on the front foot, one of them decidedly shorter than the others but complete in all its parts, and evidently capable of service in carrying a portion of the animal’s weight. The hind foot had three complete toes and a tiny remnant of a fourth which could not have been apparent externally.
As changes in the structure of the feet progressed, the central toe of the original five continued to increase in size while the adjacent digits became relatively shorter and eventually so reduced in length that they could touch the ground no longer. The smaller bones at the extremities, corresponding to the joints of our fingers and toes, eventually disappeared from the side toes. Then the longer bones of the outer digits lost the broadened supporting surface, where the missing toes had been attached, and became reduced to pointed remnants known as splints. Extreme shortening of the splint bones eventually leaves only a small knob which is often referred to as a rudimentary toe. In the skeleton of a large horse the splints are readily seen, but in some of the earlier species they are so small that they may easily be destroyed or overlooked by the collector who removes the fossilized material from the surrounding rocks. Even then, the bones of the wrist and ankle may indicate in an unmistakable manner that an additional toe once was present, for each bone is supported by another, and at the point of attachment there is a characteristic surface whose purpose is usually obvious.
Throughout the Cenozoic era the changes continued. Among the horses of the North American Oligocene were Mesohippus, approximately the size of a collie dog, and Miohippus which was slightly larger. Both were three-toed, but the rudimentary splint of a fourth toe was still present in the front foot. Parahippus and Merychippus carried on during the Miocene period, the latter being characteristic of the time, and showing, in addition to other progress, a decided trend toward the modern structure of molar teeth. There was some increase in size but the largest horse of that period was hardly more than a small pony.

A Pleistocene Horse of the Texas Plains (Equus scotti)
Hipparion and Protohippus, living during Upper Miocene and Pliocene time, represent later stages of the three-toed condition. The side toes were completely formed but greatly shortened, only the central toe touching the ground. In some of the species the outer toes had also become very slender, approaching the splint condition. By this time the molar teeth were longer and better adapted for feeding on grasses which were becoming sufficiently abundant to attract some of the forest dwellers into the open country.
During the Pliocene period, in the genus Pliohippus and also in Hipparion, the feet were far advanced in structure, with most of the species single-toed, the side digits having reached the splint stage. Pleistocene horses of the genus Equus, like living species of that genus, were strictly one-toed animals, ranging over grassy areas and highly specialized for a life in that kind of environment.
Specialization is to be noted partly in the foot and leg structures where the modifications have contributed to greater speed and travelling ability. This is of great service to an animal of the plains where food and water are often scarce, and great distances frequently have to be covered in order to obtain sustenance. The horse, as we know it, is built for speed, its limbs and feet being elongated to permit a greater stride, and also modified to decrease the weight without loss of strength. The ordinary ball-and-socket joint is replaced by a pulley-like construction which limits the direction of movement but provides an excellent mechanism for locomotion, especially over flat, open ground. Flexibility in other directions is sacrificed for greater strength, and the foot incidentally becomes less suited for other purposes.
This is what is meant by “specialization”—a departure from “generalization.” The study of fossils provides numerous illustrations of specialized development which contributes greatly to an interest in prehistoric life. Any specialized structure or habit which increases fitness for a particular way of living is also known as an “adaptation.” Quite in line with the idea of specialization and adaptation is the change which occurred in the construction of the horses’ teeth, for the dental equipment of the modern grazing animals differs widely from that of the browsing creatures which lived on the soft leaves and other plant substances of the forests.
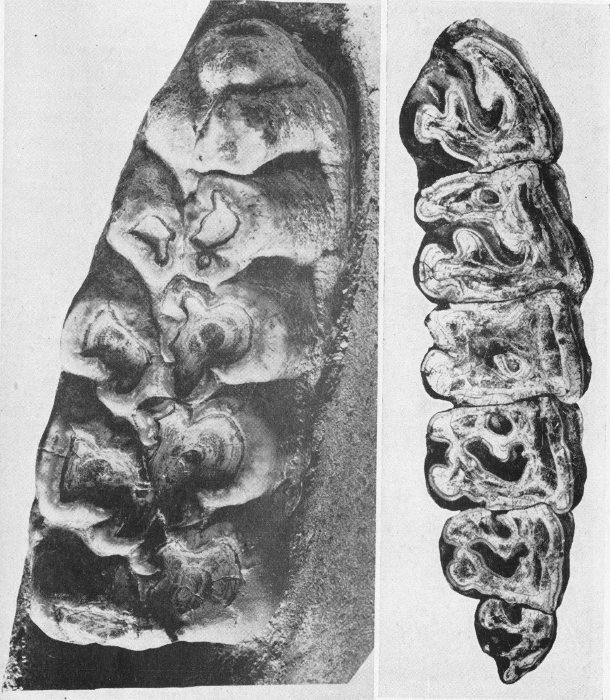
The Structure of Molar Teeth
The large lower molar of a long-jawed mastodon shows worn and unworn cusps, with the enamel layer forming a heavy border around the central dentine where the surface covering has been worn through. In the grinding teeth of rhinoceroses (illustrated at the right) the crown pattern is quite different, but both types are adapted for softer foods and are similar in having the protective enamel on the outside only. The central tooth shows the condition after the shallow surface depressions have been removed by wear.
The cheek teeth or grinding equipment of the horses underwent as complete a change as the feet. Modification resulted in a new type of tooth which enabled herbivorous animals to take advantage of a kind of vegetation which was late in arriving and has since become the principal diet of the ungulates. The grasses are coarse and harsh as compared with the leaves of forest shrubbery, requiring more thorough grinding to make them digestible. In addition they contain minute particles of silica, which is a highly abrasive mineral that quickly wears down the tooth substance, especially the softer materials found in tooth construction. An increase in the length of the tooth would offset the excessive wear but would not necessarily produce a better mechanism for grinding.
The fulfillment of the new requirements is to be seen in the change from what is known as the low-crowned, browsing type of molar, to the high-crowned, grazing type. Details of the changes that may be traced through millions of years of gradual adjustment become apparent only from the examination of a great deal of fossil material. As compared with earlier types of construction, a modern molar tooth may appear extremely complicated, but the process which brought about the improved quality is very simple. A little discussion of tooth structure, however, is required to make this clear.
A tooth, as everyone knows, is partly imbedded in the jaw, partly exposed outside the gum. In a short-crowned tooth the exposed portion is known as the crown, and the part imbedded in the jaw consists of one or more roots which are comparatively long. The crown is nearly always protected by a thin layer of hard enamel. In a grinding tooth, the working surface has a number of more or less prominent elevations known as cusps. The enamel layer completely covers this surface until wear begins. As the tooth goes into service the signs of use begin to appear; the enamel is soon worn from the tops of the cusps, and the underlying substance, called dentine, becomes exposed. This is far less resistant to wear, and as the enamel continues to be reduced the tooth becomes less efficient as a grinding device, partly because of the smoothing off of the surface, partly because of the relative softness of the inner material which is being exposed in increasing quantity. A very old molar tooth of the low-crowned type has a smooth surface from which almost the last trace of the enamel has been removed. In many prehistoric animals the enamel is of a darker color than the dentine or cement, this difference in color enabling one to see at a glance how the teeth are constructed.
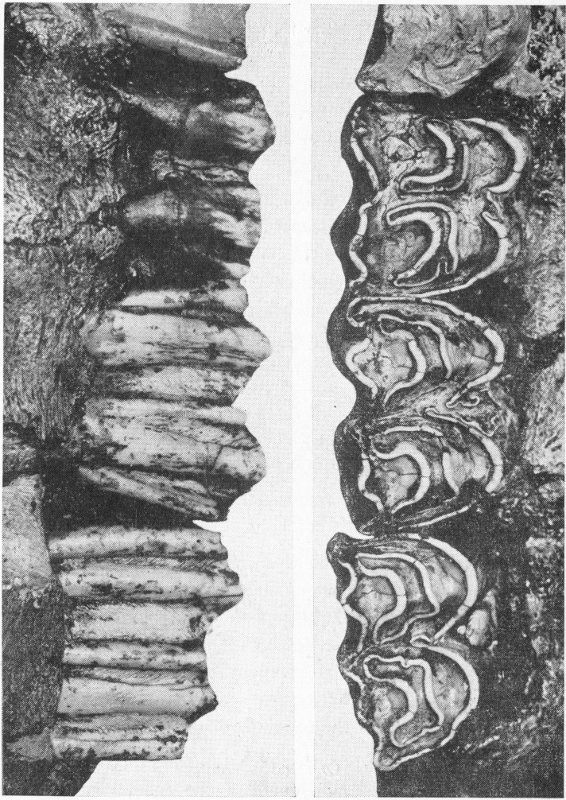
Grazing Type of Molar Teeth
The side view of the bison’s molar and premolar equipment illustrates the elongated construction which is common among grazing animals. In the pattern of the grinding surface may be seen a cross section of the enamel layers. One layer surrounds each tooth while two folded “cylinders” of the same material occupy the interior.
In a long-crowned tooth the roots are usually very short, for much of the crown itself is imbedded in jaw bone, and the longer roots are not required. Growth of the tooth is usually completed after a few years; then as it is gradually worn away it is continuously moved upward by the production of new bone under the roots, which slowly fills the bottom of the socket and continues to provide the necessary support. An equally important difference between the two types of teeth, however, is to be seen in the arrangement of the enamel, the long-crowned type being provided with this durable substance on the inside of the crown instead of having a mere protective cap on the outside.
The more complicated structure was developed from the simpler form by the easy method of deepening certain depressions located between cusps at the top of the tooth. As the crown of the tooth increased its length these depressions remained tucked in, and eventually became deep pits roughly cylindrical in shape. In addition to the enamel and dentine, a third tooth substance, known as the cement, made its appearance at about this time, and we find that quantities of this new material were deposited outside the crown enamel and also inside the enamel walls of the pit, in this way producing a firmly consolidated structure otherwise weakened by deep channels and hollow pockets. The cement differs only slightly from the dentine but is deposited while the uncut tooth is in the gum tissues of the mouth, the enamel and dentine elements being formed earlier in the embryonic tooth before it emerges from the jaw bone.
A tooth constructed by such a process, if cross-sectioned through the crown, will be found to consist of successive layers of hard and softer materials. In living animals the top of the tooth soon wears off and the enamel layers stand in higher relief because of their greater resistance to wear. A roughened surface of excellent grinding quality is thus provided, and as long as the wear continues there remains the same relative amount of enamel to retain the roughness, and resist abrasion.
Among the various types of grazing animals there is a marked difference in the arrangement and form of the enamel layers. Within a species of genus, however, the complicated enamel patterns of the molar teeth are consistently similar. In the case of horses especially, these patterns provide a most helpful key to the identification of extinct forms. The general pattern, in any of the more modern horses, may be understood more readily if the wavy enamel layers be regarded as forming a set of cylinders with deeply crinkled walls. Near the outer border of the tooth, surrounded by a thin layer of cement, is the enclosing cylinder which represents the enamel cap of the old-fashioned, low-crowned tooth. Inside of this is the central mass of dentine which has been penetrated by two of the deep pits previously mentioned. The original enamel cap has been depressed into these pits, forming two inner cylinders which are filled with cement. Instead of being circular in outline, when the cap is worn through at the grinding surface these inner cylinder walls are seen to be wrinkled and folded so as to produce a most irregular pattern. However, if several teeth of the same kind of horse are compared, it will be found that the edges of these cylinders produce figures which are remarkably uniform and characteristic for that species.
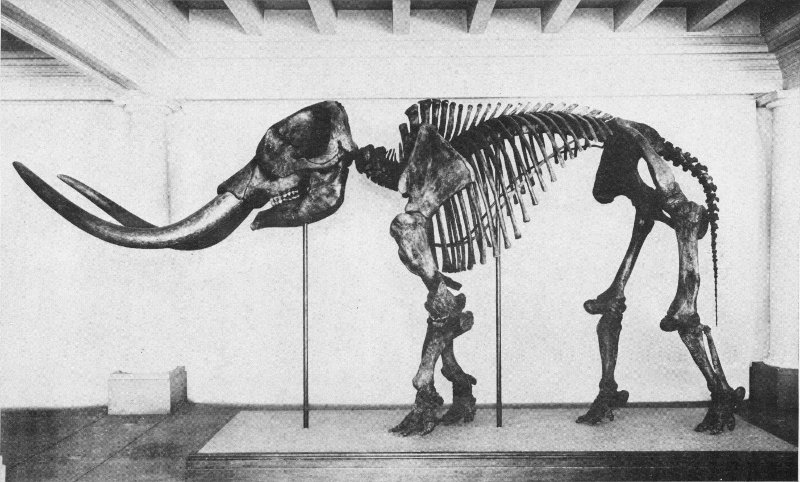
American Mastodon (Mastodon americanus)
A true mastodon of the short-jawed type.
Elephant-like mammals both living and extinct are classed together in a single order bearing the name Proboscidea. Living members of the group are the elephants, of which the large Indian and African species are best known. Among prehistoric representatives the most frequently mentioned in the popular literature of North American animals are the following:
The American Mastodon, an immigrant from Siberia which ranged over nearly all of the United States and Canada. It was principally a forest dweller, rarely found in plains regions, was abundant during the Pleistocene period and may have been known to the early American Indians;
The Woolly Mammoth, which was about nine feet tall. It ranged over British Columbia into the United States and across to the Atlantic, disappearing in late Pleistocene time;
The Columbian Mammoth, about eleven feet tall, lived in the early half of the Pleistocene period, ranging over the warmer portions of North America, including practically all of the United States and much of Mexico;
The Imperial Mammoth, reaching a height of more than thirteen feet, and becoming extinct in the Middle Pleistocene. It was a western form, remains being found from Nebraska to Mexico City.
Originally placed in the genus Elephas, the mammoths are referred to commonly as elephants, though technically they should not be regarded as such. Recent explorations and researches have added greatly to our knowledge of these animals but have also caused much confusion with regard to scientific names, for many new subdivisions of the larger group are now recognized, and it has become necessary to change some of the older nomenclature.

A Long-Jawed Mastodont (Trilophodon phippsi).
One of the Early American Proboscideans
The large mammoth exhibited by the Museum bears the impressive name of Archidiskodon meridionalis nebrascensis. Fifty years ago it might have been identified simply as a specimen of the imperial elephant and in such case would have received the old name of that species, which was Elephas imperator. But late in the last century it was proposed that the mammoths be recognized by some other name to distinguish them more sharply from living elephants. The name suggested for the new genus thus established was Archidiskodon, in recognition of the more archaic or primitive construction of the enamel plates in the mammoths molar teeth. The specific name, meridionalis, had been given to a kind of mammoth which is well known from the southern part of Europe, and the Latin name, signifying “southern,” had been applied to differentiate this species from the northern or woolly mammoth.
This mammoth, however, had disappeared from southern Europe and for many years its subsequent history remained a mystery. The late Dr. Henry Fairfield Osborn had been engaged in an extensive study of the subject, and when the nearly perfect skeleton from Angus, Nebraska, was brought to his attention he recognized it as being closely related to meridionalis, and considered it to be a record of the migration of that species into North America. Because of minor variations from the typical mammoth of southern Europe he regarded it as a variety or subspecies which had descended from the latter, and the subspecific name, nebrascensis, was added to take care of this situation, using a Latinized form of the name of the State in which the skeleton was found. With the knowledge we now have of these mammoths it becomes apparent that Archidiskodon meridionalis nebrascensis is an ancestor of the imperial mammoth, currently known as Archidiskodon imperator, and not identical with it.
This instance is typical of the manner in which prehistoric animals obtain their names. Although given a Latin form, these technical names are derived from many languages, and the root words are applied with reference to anything that happens to appeal to the author as significant. Consequently there is seldom a name of this kind which may be translated directly into natural history or science. It is a mistake to believe that these strange phrases conceal important technical information which is available only to those who are familiar with dead and foreign languages. Actually they contain nothing of the sort, and the most enlightened of the Greeks and Romans could not find it there. When a name is needed there is none better than the one provided by the specialist who is skilled in the business of naming things. Some technical ability is required, to apply the name where it properly belongs, but technical knowledge is not obtained from such sources. Names, in any form, have another purpose to serve. There is no magic in them and there need be no mystery about them.
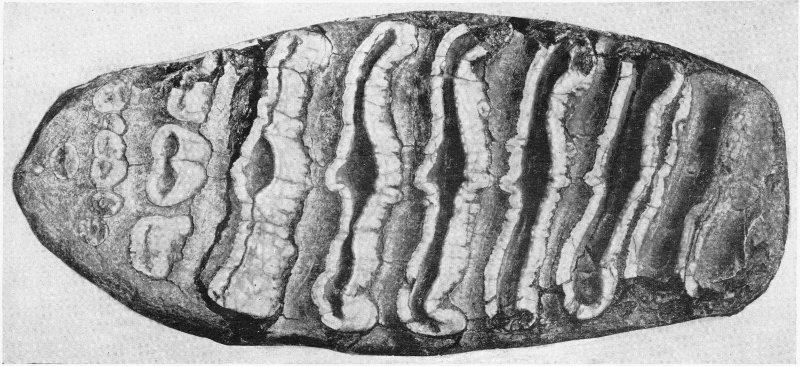
Molar Tooth of Mammoth
This type of tooth is constructed for long continued use and will withstand the wear of more abrasive foods. The position of the white enamel plates is seen in this view of the grinding surface. These plates extend all the way to the base of the tooth, which is of the long-crowned variety and not to be destroyed by the wearing away of a single outside layer of enamel.
Other specimens in the Museum collection are the long-jawed mastodonts, so named because of the elongated jaws and protruding chin which is often mistaken for a tusk. Early members of this group had more cheek teeth than later types of mastodons, and longer jaws were required for their accommodation. Some of them had flattened lower tusks which evidently were used for digging purposes. These are popularly known as “shovel tuskers.” The more modern American mastodon had shorter jaws and, like the mammoths and elephants, only one pair of tusks. Both the long-jawed and short-jawed types are represented by complete skeletons, and also by tusks, jaws, and teeth of many individuals. The American mastodons and mastodonts were of about the same size as the smaller mammoths.
The difference between mastodons and mammoths is most readily recognized in the structure of the grinding teeth, the molars and pre-molars. In the mastodon these teeth are of the short-crowned type, while in the mammoths, as in the modern elephants, they are long-crowned. The difference between these two types of molars has been described with reference to horses, and the change from the older to the modern form may be regarded as coming about in the same general way, through a series of gradual modifications. In both horse and mammoth the final development shows internal enamel extending from the grinding surface nearly to the roots. Otherwise, however, there is almost no resemblance, for the mammoth tooth is made up of flattened enamel plates, the number of which is variable for different species. In the jaws of a very young individual these plates may be seen as separate parts. As the tooth continues to grow, the plates become cemented together, and when the ends of the plate are worn down it may be observed that each consists of a layer of enamel surrounding a flat central core of dentine. The type of construction is rather more obvious in the mammoth tooth than in that of a horse, partly because of the larger size, and partly because of the relative simplicity of construction.
The earlier history of the Proboscidea is not recorded in the rocks of North America, for the group was of African origin and its migrations did not extend as far as the New World until middle Cenozoic times. The mastodons and mammoths were the largest of land animals since the Age of Reptiles, but their Old World ancestors were not conspicuous because of their bulk. Many of these ancient forms, even in the earliest stages, reveal some of the prominent characters that dominate the entire group. None of them, however, should be regarded as a miniature mammoth or mastodon, for these highly specialized types were perfected only at a comparatively recent date, and by a process that works very slowly. Among the earlier forms there were also some oddities which failed to survive or to produce a successful branch of the stock such as the elephants.

Nebraska Mammoth
(Archidiskodon meridionalis nebrascensis)
The earliest known member of the order was Moeritherium, an animal of the size of a tapir, living in Egypt during the late Eocene and early Oligocene time. At this stage the characteristic specializations leading to the mastodons and mammoths were apparent but not far advanced. The proboscis was probably much like the flexible snout of modern tapirs, for the need of a long trunk had not yet arrived. In upper and lower jaws the second pair of incisor teeth were becoming large and prominent. The enormous tusks of the mammoths later developed from the enlargement of the same pair of upper incisors, and in some of the long-jawed mastodonts the lower pair also produced large tusks, though frequently the lower tusks were not prominent.
Dinotherium had downward-growing tusks in the lower jaws, none in the upper. This genus was fairly common in the Miocene of Europe, Asia, and Africa. In the tropics it survived throughout the Pliocene and possibly into the Pleistocene. Some of the species acquired the size of elephants, but it is apparent that they were not ancestral to any of the more progressive types. They are to be regarded rather as an offshoot from the main line of descent.
In 1859 only ten species of the elephant-like mammals were known, and all were referred to a single genus. At the present time eleven genera appear to be well founded, and the number of recognized species has reached a hundred, if it has not already passed that figure. New discoveries are expected to add to the existing total. With this mass of material before us we note certain definite trends among the more progressive types. The increasing weight was accompanied by the development of strong, upright limbs in which the bones have a columnar position instead of the angular assembly which prevails among most of the mammals. As the tusks increased in size there was a shortening of both skull and neck to bring the weight closer to the point of support. The front teeth disappeared except the second pair of upper incisors which remain as tusks in the modern elephant. The cheek teeth present in the shortened jaws of the mammoth were reduced to one pair at a time in the upper set and another pair below. From a simple, low-crowned origin these grinding teeth developed into the more successful high-crowned pattern with numerous plates of enamel inside. A prehensile upper lip acquired the length and usefulness of the elephants trunk.

Rancho la Brea Fossils
One of the most unusual of the many animals that have been taken from the tar pits is the large ground sloth, seen at the left in this group. Such sloths were very abundant during Pleistocene time, and some may have lived up to a few thousand years ago. Archaeologists have found indications that these creatures may have been hunted by cave-dwelling peoples of the American Southwest.
Other skeletons include the saber-tooth tiger, characterized by the long curved upper canine teeth which undoubtedly were used for stabbing and slashing, and the dire wolf, the smaller of the two which are facing the sloth. The artist’s reconstruction of this scene also shows the great vulture, Teratornis, which is the largest known bird of flight.
Over-specialization in the production of tusks appears to have been the principal factor in the downfall of the mammoths. The large size of the animals and the difficulties of finding sufficient food to sustain life must have been a serious handicap at times, but their ability and inclination to travel over long distances enabled some of them to find tolerable living conditions until the end of the Glacial Period. They are now extinct and the nearest living relatives are the elephants, somewhat reduced in size of tusks and body but otherwise very similar.
There are many other tribes of mammals whose ancient history is partially known though broken by periods of time for which there is no fossil evidence. All have undergone changes in which various forms and degrees of specialization are featured; this general process is best revealed by the horses and elephant-like animals which have left a clearer record. For other groups the story would differ but little except as to names and specific details.
The La Brea tar pits, as they are often called, provide a remarkable record of Pleistocene life in southwestern North America. Scattered over an area of about thirty acres just off Wilshire Boulevard in Los Angeles, these bone deposits were known, as far back as 1875, to contain the remains of prehistoric animals. It was not until 1905, however, that their value was recognized by paleontologists. In that year the University of California began an investigation, and excavations were carried on at intervals by various institutions during the next ten years. A great deal of material was acquired by the Los Angeles Museum of History, Science, and Art, where many skeletons, skulls, and other interesting specimens have been placed on exhibition.
The pits have the form of small craters formed by the seeping of oil from the underlying rocks. The seeps appear to have been active during part of the Pleistocene period but apparently not at the beginning. The oil is rich in asphalt which has served as a preservative for the bones, and owing to its sticky properties has been an effective animal trap for thousands of years.
The fossil beds at present are of oil-soaked earth and sand. In past times there must have been a greater percentage of oil, often concealed by a layer of dust or pools of water. The large number of carnivorous animals found in the deposits suggests that they were attracted by the cries and struggles of creatures wandering carelessly into the asphalt and serving as live bait to keep the traps in continuous operation.
Animals found there include many species still living in the locality, some that have migrated to other territory, and a large number that have become extinct. Among the latter may be mentioned species that differ but slightly from living relatives, others that have left no descendants. 89 Horses, bison, and wolves, though extinct species, were of relatively modern types. On the other hand the large sloths and saber-tooth cats seem rather out of place. True cats are represented by the mountain-lion, bob-cat, and a species of lion which is nearly one-fourth larger than any of the great cats of the Old World. A long-legged camel, with a height of approximately eight feet to the top of the head, was among the native animals of the district. Skunks, weasels, badgers, squirrels, rabbits, bear, deer, and antelope were more or less abundant.
The La Brea group exhibited by the Denver Museum of Natural History includes the following species: horse (Equus occidentalis), bison (Bison antiquus), wolf (Aenocyon dirus), saber tooth (Smilodon californicus), sloth (Mylodon harlani). Horses had entirely disappeared from the North American continent by the time the first white man arrived. Equus occidentalis was one of the several species living during the Pleistocene period, this one apparently being restricted to California and perhaps adjacent states. Bison antiquus was slightly larger than the plains bison of recent times and had it horns set at a characteristic different angle. The species was first described from Kentucky and appears to have had a wide distribution.
The wolves in this group are about the size of timber wolves, but have heavier skulls with less brain capacity, massive teeth especially adapted to biting and crushing large bones, and limbs of rather light construction. They probably assembled in packs where meat was abundant and, hunting in this fashion, were able to attack and overcome the larger ungulates and edentates. To most visitors the large ground sloth is the most interesting animal of the group. This edentate animal is shown at the edge of the pool with one foot stuck in the “tar.”
The edentates are a group of primitive animals with very simple teeth, if any. Teeth are usually lacking in the front part of the mouth, sometimes entirely absent, as among anteaters. Better known living representatives of the group are the tree sloths, armadillos, and anteaters of South America. Ground-sloths were prominent among South American mammals during much of Cenozoic time. During Pliocene and Miocene time there was a marked tendency to large size, and it was principally during these two periods that they appeared in the United States area.
Mylodon was one of the larger North American ground-sloths. Its teeth, without the protective enamel which is present among higher mammals, are restricted to the cheek region, and have the form of simple pegs; instead of being specialized they stand close to the extreme of generalization. The construction of the entire skeleton is massive, suggesting great strength with slow movements. The hands are well developed, provided with stout claws, and must have served the creature well as protection against attacks by predatory neighbors. We have some idea as to what caused the extermination of the ground-sloths in this particular region, but the complete disappearance of such a large and widely distributed group at the close of the Pleistocene period is a mystery that may never be explained.
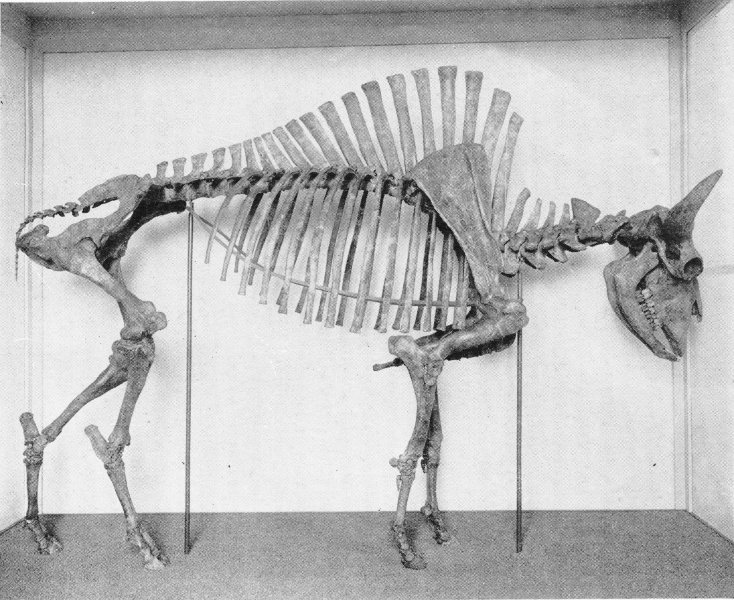
The Folsom, New Mexico, Bison (Bison taylori)
The saber-tooth cat, sometimes referred to as a tiger, was specialized as a meat eater though hardly as a hunting animal. In the La Brea region its principal food was probably the flesh of the sluggish ground-sloths. The size was equal to that of the African lion, with hind limbs slightly longer and the front legs more powerfully developed. The most remarkable characteristic is to be found in the development of the upper canine teeth and modifications of the skull which were necessary to enable the animal to use these teeth as weapons.
In order to make the “sabers” effective it was necessary to get the lower jaws out of the way, and this was provided for in an unusual type of hinge which enabled the mouth to open wider than is possible in the case of the less specialized carnivores. Judging by all the structural features of the skeleton, Smilodon could not have lived well on small animals, for it was not equipped to capture that kind of prey. It is evident that large mammals were preferred, and that the method of attack was to spring upon the victim and cling there with the powerfully developed fore limbs until the kill was completed by stabbing into a vulnerable spot. That the position of the large sabers near the front of the mouth interfered with normal feeding, is a reasonable conclusion. There are also anatomical features which lead to the belief that this carnivore was a blood sucker, perhaps more than it was meat-eater.
If most of these conclusions are correct we have here another case of over-specialization and a possible explanation of the extinction of two species. Such evidence as we have is far from conclusive, for there is no proof that Rancho La Brea was the last stand of either the saber-tooth or the ground-sloth. Both races were widely distributed and their living conditions could not have been exactly duplicated in other localities. It has been suggested, however, that Smilodon ate the last of Mylodon, and starved soon afterward because it had become unable to partake of other foods. The conjecture is offered for what it is worth, together with the facts on which the story has been based.
The geological record for Pleistocene time is not as complete as one might imagine. Numerous localities have produced representative fossils but the yield is rarely large enough to solve many of the riddles which are constantly arising as investigation proceeds. Aside from those areas which bordered the retreating ice cap and where living conditions were far from favorable, the sedimentary deposits of this period are not continuous over large areas. Many Pleistocene fossils are found in stream channel beds which are always subject to removal by subsequent floods.

Early Man in North America
There is abundant evidence to indicate that the great elephants of Pleistocene time were hunted by primitive Americans whose only weapons were darts or spears tipped with points of stone. A skull and the lower jaws of several mammoths are shown here.
Isolated patches of fossil-bearing sediments frequently record the migration of animals in unmistakable terms, but the details of the wanderings and the conditions encountered in the newly established habitats are often left in doubt. To correlate the facts revealed at one locality with findings at other places and, if possible, to date all prehistoric events with a greater degree of accuracy are among the major tasks of current investigations.
The Pleistocene or “Ice Age,” and the Recent period in which we are living at the present moment are not sharply separated by any event readily recognized or dated, and the two combined are of very short duration as compared with other periods more clearly established by the passing of centuries. Together they comprise the Age of Man as commonly recognized, with about a million years representing the Pleistocene period, some ten to twenty thousand years the Recent. When geologists of the nineteenth century suggested that the coming of man should be regarded as the beginning of a new era, the name Psychozoic was proposed, and to some extent this term has been applied to the present period. More in keeping with other period names is Holocene, meaning entirely recent. Common usage, however, applies the simple term Recent to this unfinished chapter which is also without a clear-cut beginning.
Zoologically, man is merely one of the creatures that arrived in the course of time, along with other mammals. Just when he arrived and how he looked at the time of his coming cannot be determined from a study of fossils. Perhaps it is of no importance. There is nothing to indicate his existence before the Cenozoic, no completely satisfactory proof of existence before the Pleistocene period. As with other inhabitants of the earth, it is probable that he became prominent only after a great deal of competition with other creatures which kept his ancestors submerged for thousands of years. The Ice Age, with its check upon the progress of competing animals, undoubtedly gave him an advantage. His superior mentality enabled him to overcome adversity by methods not available to other mammals; his inventive and mechanical genius must have been greatly strengthened by his experience during this interval.
At about this point, where prehistory begins to merge into history, 94 the geologist and paleontologist must let other interpreters carry on. Archeologists and anthropologists take up the work, and through their efforts many details have been added to our knowledge of the human race. The study of biology, which is the science of life, has provided an instructive viewpoint that enables us to see ourselves against the vast background built up by investigations into the nature of the earth and its ancient inhabitants. This science deals with living creatures as organisms—plants and animals so organized as to be capable of existence only in an environment which provides exact life requirements.
The Age of Man has been variously characterized as an age of soul, of higher intelligence, of culture, and finally, of civilization, freedom and democracy. The “crowning glory” of the organic world is pictured in history as a creature who has busied himself for thousands of years with the building up and tearing down of civilizations. Prehistory reveals this habit as something unique in the human character, for there is no other organism that has specialized so persistently in the creation of its own environment, no other that has had the combined power and talent to produce so much change.
More than anything else, the prehistoric record is a lesson in adaptation, which in its broadest sense means fitness for life under particular conditions, and always subject to organic law. Man’s efforts to bring about an adjustment between himself and his civilization have centered largely on the method of forcing himself into the mold that happens to be present, one pattern today, another tomorrow. No creature of the past has had to adapt itself to anything so radically new or so thoroughly revolutionary. The vital problem now is whether this man-made environment will prove helpful or disastrous.
Though one of its names is “culture,” it has grown sporadically and unevenly, with little evidence of the cultivation that is implied and required. Parts have been expanded to extraordinary proportions while others equally essential have been retarded in their growth. A more intelligent handling of this environment factor seems to be possible, and the present mania for “organization” may become tempered with an awakening consciousness of organic requirements where organism and environment are involved. Once we grasp the idea that “culture” results from man’s effort to improve his living, by putting into his environment something that was not there before—then, surely, this history of a billion years of living, and as many “ways of life,” should teach us something we ought to know as we go into an all-out endeavor to teach a whole world how to obtain a one-and-only way.
We may stand at the beginning of an era for which an appropriate 95 name has not yet been suggested. Civilization, on the other hand, may provide only a minor epoch to be added in some remote time to the story of fossils.
The literature pertaining to fossils is widely scattered and usually too technical for the layman. It is better to use the resources of the nearest library than to feel that a specified list of books is necessary.
Any textbook on geology, zoology, or botany will provide helpful information. Most books of this type will be found interesting and readable if used to solve definite problems suggested by the student’s immediate curiosity. Very few can be read from beginning to end without a great deal of effort and discouragement.
The following have been prominent among the books consulted by the author:
Textbook of Geology; by Pirsson and Schuchert. This work has undergone several revisions and currently appears in two volumes: Physical Geology by Longwell, Knopf, and Flint; Historical Geology by C. O. Dunbar. Published by John Wiley & Sons. (Historical geology covers the entire range of prehistoric life—plant, invertebrate, and vertebrate.)
Historical Geology (The Geologic History of North America); by Russell C. Hussey. Published by McGraw-Hill. Concise, interesting, and informative.
Geology and Natural Resources of Colorado; by R. D. George. Published by the University of Colorado. Contains an excellent summary of the historical geology and sedimentary formations of Colorado.
Vertebrate Paleontology; by Alfred Sherwood Romer. Published by the University of Chicago Press. This is one of the most comprehensive and up-to-date treatments of the subject for students desiring to go beyond the elementary stage.
A History of Land Mammals in the Western Hemisphere; by William Berryman Scott. Published by The Macmillan Company. This well-known account of living and extinct mammals is one of the favorites among students.
The Age of Mammals; by Henry Fairfield Osborn. A classic in this field of literature, but for advanced reading. The book is now out of print.
The Dinosaur Book; by Edwin H. Colbert. Published by the American Museum of Natural History, New York. An illustrated story of amphibian and reptilian evolution.
Down to Earth; by Carey Croneis and William C. Krumbein. Published by the University of Chicago Press. An excellent popularization of the earth sciences—geology and paleontology.
Lexicon of Geologic Names of the United States; compiled by M. Grace Wilmarth. Bulletin 896 (in two parts) of the United States Geological Survey. A rich source of information concerning the age, character, and distribution of geologic formations, with numerous references to fossil-bearing beds.
Bibliography of North American Geology (including paleontology); various bulletins of the United States Geological Survey. Where library facilities provide access to the technical literature of museums, universities, and scientific societies, this is a valuable aid in locating publications dealing with original work in paleontology. Bulletins 746 and 747 cover the years between 1785 and 1918; Bul. 823 (1918-1928); Bul. 937 (1929-1939); Bul. 938 (1940-1941); Bul. 949 (1942-1943); Bul. 952 (1944-1945); Bul. 958 (1940-1947); Bul. 968 (1948); Bul 977 (1949). Preparation is a continuous process with recent bulletins appearing at one or two year intervals.
Ancient Man in North America and Prehistoric Indians of the Southwest; by H. M. Wormington. Published by Denver Museum of Natural History, City Park, Denver 6, Colorado. Both volumes contain authentic and up-to-date accounts of early American cultures.
Geologic Maps. United States Geological Survey: map of the United States (1932); map of Colorado (1935). Geologic maps of a few other states are available; information regarding these may be obtained from state universities or state geological surveys.
Note: Bulletins of the U.S.G.S. are purchasable from the Superintendent of Documents, Washington, D. C. Maps are sold by the Director of the Geological Survey, Washington, D. C.